【www.zhangdahai.com--其他范文】
马锦征 苏超 杨洁 魏民新
300070 天津医科大学基础医学院生物化学与分子生物学系(马锦征、杨洁);
518057 深圳,香港大学深圳医院心血管医学中心(苏超、魏民新)
随着我国国民生活方式和年龄结构的巨大改变以及人口老龄化的加速发展,老年易患疾病的发病率达到了前所未有的高度。目前,我国心血管病患病率处于持续上升阶段,据推算心血管病现患人数3.3亿,其中冠心病1 139万、心力衰竭890万、先天性心脏病200万、高血压2.45亿[1]。2018年,我国心血管病死亡率仍居首位,农村心脏病死亡率为162.12/10万,城市为146.34/10万[1]。心脏疾病负担日渐加重,已成为我国重大的公共卫生问题,防治刻不容缓。
心脏作为高耗能器官,静息状态下需消耗人体10%的能量来维持正常功能,每日产生 6 kg ATP,几乎是心脏质量的3倍。为产生足够的能量,心肌细胞对能量底物的选择多样而灵活,其中包括糖类、脂肪酸、氨基酸、酮体和乳酸等,可随血液循环中底物浓度和含氧量不同变化进行相应调整,这一行为利于心脏处于饥饿、压力超负荷或氧化应激等不利状态下确保能量供应,维持功能稳定。
正常状态下,成熟的心肌细胞中脂肪酸的β-氧化占ATP供能的80%,葡萄糖氧化占12%,乳酸氧化及糖酵解分别占7%和1%,可见ATP供能主要来源于脂肪酸β-氧化[2]。正常心脏中,来自脂肪酸的脂酰辅酶A和来自糖类的丙酮酸是线粒体的主要能源[3-4]。为了维持有氧能量代谢,有氧ATP产生的主要场所线粒体,在心肌细胞内占据30%的体积。心肌细胞通过不同底物产生的小分子代谢产物也可以在代谢相关信号通路中发挥作用,转而调节心肌细胞微环境,影响心脏功能[2, 3, 5]。
1.1 心脏发育过程中的代谢方式转变
胎儿心脏作为胚胎发育过程中最早形成的功能器官,一般在孕23 d开始跳动,4周左右开始行使泵血功能。由于循环血中游离脂肪酸含量非常低,胎儿心肌主要以葡萄糖和乳酸作为ATP的主要来源。胎儿心肌细胞微环境中含有较高水平的葡萄糖和产物乳酸,有利于连续分裂细胞的酯膜组分、氨基酸、核酸等生物大分子的合成。这种以无氧糖酵解为主要能量来源方式的情况,同时影响了糖代谢和脂肪酸代谢通路。胎儿循环血中乳酸含量较成人高,心肌细胞表面涉及糖酵解的酶分子高度表达,主要有葡萄糖转运蛋白1、己糖激酶1等,而涉及脂肪酸代谢及线粒体合成的分子表达维持在较低水平。低氧环境有助于维持多能细胞未分化状态,随着细胞分化,主要能量来源由无氧酵解转为氧化磷酸化[6-7]。
无氧糖酵解的降低和脂肪酸β-氧化的升高同时发生在心肌的成熟过程中[6]。刚出生的新生儿心肌短期脂肪酸氧化效率依然不高,但在接下来的几天内会出现剧烈增长。多能胚胎干细胞的线粒体不成熟,主要分布于核周且无网络状结构;
而完全分化细胞有足够多结构完整的线粒体,以管状网络结构均匀分布在胞浆。成熟线粒体内存在丰富的氧化磷酸化,线粒体膜电位上升,耗氧量及ATP产出都有所提高。而随着线粒体成熟,电子传递呼吸链复合物酶及三羧酸循环酶表达量上升,而糖酵解酶表达降低。
1.2 心肌生理性增厚过程中的代谢方式转变
静息状态下,健康的心脏为了维持全身血供需要机体总耗氧量的10%;
而剧烈运动时,这一消耗量可达到之前的10倍。由于心肌细胞对代谢底物的选择有极高的灵活性,可支持短时间高耗能的剧烈跳动。而面对长期高耗能的需求,心脏会发生一系列适应性形变,包括心肌细胞的增生、心室壁的增厚、心腔的扩大等,从而达到更高的每搏射血分数和左室收缩能力。早期的病理性心肌肥大心脏形态变化与生理性心肌增长相似,但代谢旺盛产生更多的能量这一特点是生理性肥大特有的[8]。
1.2.1 运动诱导的心肌增厚 适量运动有助于保持健康、延长寿命,尤其在慢性心血管病患者中,定期有氧运动对其病情起极有效的控制作用[8-9]。有氧运动如跑步主要促进每搏输出量的增加,而举重类无氧运动对心血管的影响主要反映在压力负荷耐受性的增强。由于终末分化心肌细胞失去了增殖能力,所以不论何种运动方式均促进心肌细胞体积的变大,而非细胞数目的增多[10]。
生理性与病理性心肌肥大从转录水平就有明显差异,病理性肥大心肌中出现心肌“胚胎化”现象,胎儿心脏特异的肌球蛋白重链β在成年心肌中出现,且心房利钠肽/B型利钠肽表达上升,这些现象在生理性肥大心肌中均未出现。生理性肥大心肌中线粒体生物合成相关基因表达上调,同时包括电子传递链、脂肪酸氧化相关酶等一系列线粒体有氧氧化蛋白表达量上升,是心肌面对增大的能量需求时做出的适应性变化。
在运动过程中骨骼肌和心肌中,过氧化物酶体增殖物激活受体γ共激活因子1α(peroxisome proliferator-activated receptor γ coactivator-1α,PGC-1α)表达显著增多[8],运动诱导产生的PGC-1α促进线粒体生物合成诱导线粒体基因mtDNA的合成,并促进脂肪酸氧化磷酸化过程。过氧化物酶体增殖物激活受体α(peroxisome proliferator-activated receptor alpha,PPAR-α)和PGC-1α的结合保护心肌不被过氧化物损伤,延缓了心力衰竭病程;
同时调控心肌脂肪酸摄取和氧化。CBP结合转化激活因子4被证实在运动适应性肥大心肌中促进细胞生长和增殖,同样可保护心肌不受缺血/再灌注损伤。而在所有细胞内信号通路中,与心肌生长最密切相关的是PI3K/Akt信号通路[11]。其失活或突变会导致心肌损伤和收缩能力下降,在上游基因过表达和运动诱导通路激活的情况下均对心肌起到一定的保护作用[10-11]。
1.2.2 妊娠诱导的心脏重构 与运动一样,妊娠会促进心脏质量的可逆增加。早期研究结果表明,妊娠会增加每搏输出量和心率。妊娠期间心肌肥厚的程度可能超过运动时通常发生的程度,左心室质量增加多达50%;
然而,由于妊娠晚期体重大幅增加,母体心脏质量与体重之比实际上降低了,而妊娠晚期甚至可能增加心肌缺血的危险。尽管如此,妊娠期间的轻度心脏功能障碍通常并无太大危害,因为它会在产后逆转。妊娠期血管生成增加,而并不伴随心肌纤维化,有助于心脏适应妊娠这种特殊的生理形式。
在代谢方面,妊娠可轻微降低循环葡萄糖水平,但同时酮体、非酯化游离脂肪酸、乳酸和三酰甘油的水平较高。激素水平和环境因素共同作用下促进心肌代谢重构,以增加脂肪分解代谢为代价来节省葡萄糖。其中黄体酮增加丙酮酸脱氢酶激酶4(pyruvate dehydrogenase kinase isozyme 4,PDK4)的表达,导致葡萄糖氧化的抑制和脂肪氧化速率的提高[12];
而妊娠期间成纤维细胞生长因子21水平升高也会诱导PDK4并增加心脏中的脂肪酸氧化[13]。而内皮一氧化氮合酶,通常在运动中被上调或激活,可激活脂肪酸氧化并抑制妊娠犬类心脏中的葡萄糖氧化[13]。
妊娠期间心肌葡萄糖分解的减少对心脏生理性发育十分重要。二氯乙酸可降低PDK4介导的葡萄糖氧化抑制,从而防止妊娠引起的心脏增长。为了维持心脏生长所需的葡萄糖,心肌选择提高脂肪酸氧化供能的比重。妊娠诱导的心脏生长会伴随葡萄糖氧化减少这一现象,与在运动背景下的发现一致,其中葡萄糖分解代谢的间歇性减少均驱动了生理生长所需的代谢方式的转化。不论在妊娠期间还是运动诱导,这一过程常常是通过成纤维细胞生长因子21诱发的[2]。尽管葡萄糖分解代谢的减少可能会为肌细胞生长所需的合成代谢反应腾出葡萄糖,但生物合成途径如何随着妊娠而变化仍未得到研究。此外,在妊娠诱导的心脏生长的背景下,代谢在表观遗传重编程、代谢物信号传导和氧化还原平衡中的作用仍不清楚(图1)。
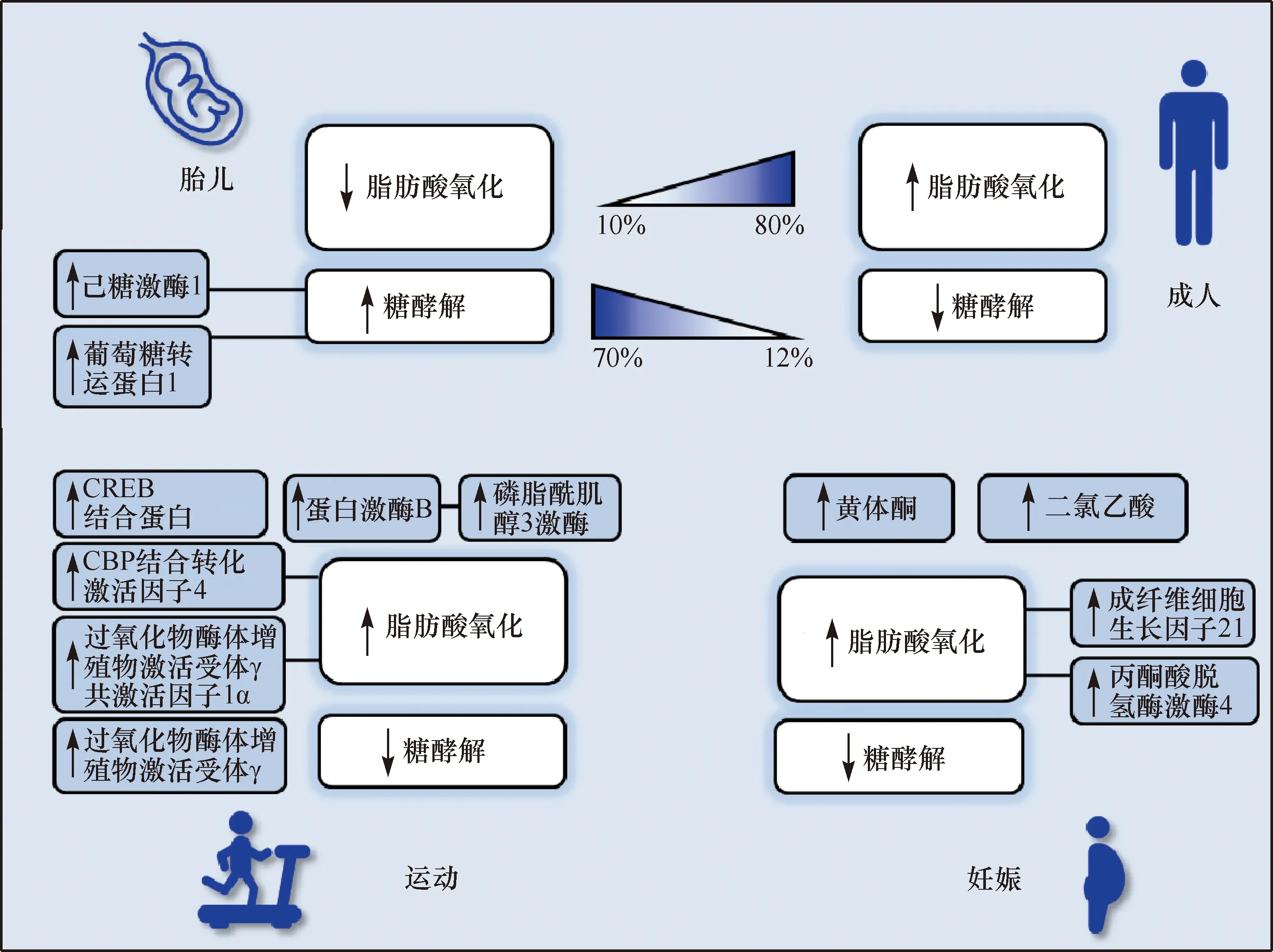
图1 生理情况下心肌发育、运动和妊娠引发的代谢转变
2.1 缺血性心肌病
缺血性心肌病主要指心肌由于缺血缺氧,导致心肌纤维化和心肌梗死症状单独或相互组合出现为主要特征的一组临床症候群。多由冠状动脉粥样硬化为主要病因,进一步发展为严重心功能不全。
正常心肌以脂肪酸β-氧化为主要供能方式,但缺血情况下丙二酰辅酶A含量减少,线粒体对脂肪酸的摄入不足。而心肌糖酵解产能速率远低于有氧氧化,同时产生的大量质子和乳酸,破坏原有的离子稳态[14]。线粒体膜电位的稳定对细胞器稳态及呼吸链电子传递至关重要,长期失衡导致线粒体ATP生成下降。
对能源物质利用的灵活性可保护梗死区的心肌细胞[15]。有研究证实,支链氨基酸在正常心肌中抑制丙酮酸激酶活性,降低糖类摄取及有氧和无氧氧化,同时促进脂肪酸代谢[16]。支链氨基酸这一作用在应激状态下保护心肌细胞不被诱导凋亡[15-16]。
缺血再灌注后,堆积的脂肪酸充分氧化,使葡萄糖氧化过程被抑制。但再灌注突然补充的大量氧气和营养,同时产生大量活性氧因子,造成氧化损伤甚至细胞死亡[14, 17]。代谢组学显示,活性氧造成的氧化损伤与琥珀酸有关,缺血区域琥珀酸堆积,其浓度越高,活性氧损伤越严重;
对梗死区琥珀酸浓度的控制存在一定治疗意义[18-19]。
2.2 心肌肥大、扩张型心肌病
肥厚型心肌病以心室肥大、室间隔非对称性肥厚、舒张期充盈异常及左心室流出道受阻为特征。病因常为多种造成心脏泵血后负荷加重的疾病,长此以往引发代偿性肥厚。如果合并动脉粥样硬化可进一步导致心肌供血不足,加速心力衰竭[20]。
作为慢性进行性病理变化,心肌肥大带来的代谢方式转变也是缓慢发生的。压力超负荷性肥大心肌中,长链脂肪酸氧化和糖类有氧氧化均下降,糖酵解速率反而上升,这种情况也称作心肌细胞“胚胎化”。在肥厚心肌中糖酵解供能可以达到19%,而脂肪酸降低至55%左右[21]。在动物模型中,经主动脉缩窄术后1 d小鼠和对照组代谢组学差异不明显,但约1~8周后手术组和对照组小鼠代谢产物变化明显,且随时间进行性增大[18]。8周后心肌细胞中丙酮酸及脂代谢产物均明显下降,其中鞘磷脂含量的降低非常明显,可能是由于受损心肌中鞘磷脂合成过程中中间产物的减少[22]。鞘磷脂被认为在心肌细胞的增长和自噬过程中发挥重要作用[18, 23]。
各类研究中普遍认为肥大心肌中脂类氧化产能下降,转而以糖酵解为主要供能方式。也有研究表明,肥大心肌中脂肪酸氧化速率和产能其实有所提高,然而线粒体对脂类的摄取和转运环节出现问题,导致重要中间产物酰基肉碱含量下降,使代谢底物不足以供应受损心肌细胞巨大的能量需求。在自发高血压模型大鼠中脂肪酸转位酶(FAT/CD36)突变,出现进行性心肌肥大,心肌糖酵解速率升高,脂肪酸摄取明显缺陷,而循环脂肪酸含量有所增加[24-25]。
以往的研究结果中普遍显示肥大和衰竭心肌中糖类代谢速率升高,糖酵解产物丙酮酸含量有下降,与乳酸含量的升高相符,受损心肌的乳酸/丙酮酸比例会达到正常心肌的2.5倍,显示了无氧代谢供能比重的增加[18]。PDK4作为丙酮酸脱氢酶(pyruvate dehydrogenase,PDH)最重要的激酶,其表达量直接影响了丙酮酸生成乙酰辅酶A的反应[26]。肥大心肌中长链脂肪酸氧来源的丙酮酸含量降低,糖类有氧氧化占主要地位[18]。三羧酸循环中苹果酸-丙酮酸的转化在肥大心肌增加,丙酮酸含量降低[18]。受损心肌中磷酸戊糖途径降低,可能是受心肌细胞氧化应激影响。
病程进一步延长的大鼠体内可出现糖酵解速率下降,心肌细胞出现胰岛素抵抗和代谢重构的现象[27]。受到胰岛素刺激的细胞通过胰岛素结合受体通路引发下游一系列蛋白磷酸化反应,使贮存在细胞浆囊泡内的葡萄糖转运蛋白4转位到肌细胞膜表面,行使葡萄糖转运功能[28-30]。
2.3 糖尿病心脏病
糖尿病患者长期处于高血糖状态,导致各种组织,特别是眼、肾、心脏、血管、神经等的慢性损害和功能障碍。糖尿病患者发生心力衰竭的概率约为22%,最终表现出心肌纤维化、肥大、微循环障碍和基膜增厚等症状[31]。
糖尿病患者血糖异常导致线粒体功能缺陷可能是心血管疾病发生的原因。在肥胖或高血糖患者和动物模型中,葡萄糖氧化速率下降,脂肪酸成为这类患者心肌几乎唯一的能量代谢来源[32]。前文提到,心肌对代谢底物的选择非常灵活,其代谢方式也相对复杂,正常情况下心脏同时利用脂肪酸和葡萄糖氧化供能。过高的脂肪酸摄取和使用,与心肌细胞的胰岛素抵抗有关,进一步导致心肌肥大,心肌功能受损[33]。
发生胰岛素抵抗的心肌细胞中,脂肪酸转运分子发生从胞质到胞膜的迁移,脂肪酸摄取因此发生剧烈升高。部分蛋白组学研究也证实了脂肪酸转运相关蛋白,如脂肪酸结合蛋白、长链脂肪酸合成酶等,与糖尿病患者进行性心肌病相关。除此之外,脂肪酸作为PPAR家族的一种连接体,其含量的升高引起PPAR家族调控的一部分基因表达上升,其中就包括CD36、肉毒碱棕榈酰转移酶1和长链脂肪酸脱氢酶1,导致心肌线粒体脂肪酸氧化速率加快。同时PPAR-α通过PDK4促进PDH磷酸化,使其由胞膜位移至胞浆,抑制了糖酵解过程[34]。
2.4 心力衰竭
前文提及,在正常心脏中约60%~90%的能量来自脂肪酸氧化,其他部分由葡萄糖、氨基酸和酮体等补足[35]。目前看来,心肌肥大与终末期心力衰竭在代谢相关基因的表达水平上表现类似,但肥大心肌代谢组学和正常心脏差异并不大[2, 35]。心力衰竭病程进一步发展,受损心肌细胞线粒体功能缺陷,严重影响有氧氧化速率,此时会出现严重的ATP缺口。在终末心力衰竭患者的代谢组学研究中,支链氨基酸代谢对糖酵解的影响以及酮体的重要作用逐步被重视[3, 15]。患者脂肪酸有氧氧化速率降低,而同为脂类的酮体氧化相关酶的水平提高[36]。在心力衰竭过程中,心肌代谢病理状态会引起一系列全身反应,而血液循环中这些代谢产物也会反作用于心肌[37](图2)。

图2 心脏疾病引发的代谢改变
代谢通路的转变在心脏疾病的发生发展中占有突出地位,针对各类疾病中代谢相关靶点的治疗方法也取得了巨大进展。目前临床利用代谢调节的方式对缺血性心肌病进行治疗的原理,主要是诱导心肌从游离脂肪酸向主要利用葡萄糖的转变,以增加每单位耗氧量的ATP生成。此类药物中以曲美他嗪、雷诺嗪和哌克昔林应用最多,证据也最充分[38]。恩格列净是一种新开发的抗糖尿病药物,通过高度选择性地抑制钠-葡萄糖共转运蛋白2来降低高血糖症。有证据表明,恩格列净对缺血后出现急性心肌功能障碍的非糖尿病大鼠的心功能有保护作用,其作用机制与降低葡萄糖水平和增加酮体氧化代谢有关[39]。
糖尿病作为代谢紊乱类疾病,针对糖尿病心脏病的代谢治疗方法较多。胰高血糖素样肽1(glucagon-like peptide-1,GLP-1)可在离体小鼠心脏中通过增加葡萄糖摄取及冠状动脉血流来保护心脏[40]。研究显示,GLP-1与心肌细胞表面GLP-1受体结合,上调心肌细胞的葡萄糖转运体4,从而加强心肌细胞摄取葡萄糖,改善缺血心肌的葡萄糖代谢,最终改善心功能[41]。最新对于线粒体呼吸的辅酶Q10的研究表明,其可减少糖尿病小鼠的氧化应激[42],还可通过防止活性氧的增加来减轻动物模型中的氧化应激和心脏功能障碍[43]。
心力衰竭慢性代谢转变中,线粒体损伤的缓解和线粒体功能的逆转和恢复在代谢疗法用于心力衰竭治疗的过程中至关重要。线粒体脱乙酰酶Sirtuin-3通过阻止关键的线粒体酶乙酰化和维持线粒体功能的活性改变,并通过增强对心脏能量代谢至关重要的PDH和ATP合成酶的乙酰化水平,抑制其活性[44]。有报道称,甲状腺激素刺激心脏线粒体生物发生,增加心肌线粒体质量、氧化磷酸化、酶活性、线粒体蛋白质合成和mtDNA含量,在小鼠模型中有效延缓了心力衰竭进程[45]。梗死后心力衰竭与葡萄糖转运蛋白的上调有关,而在小鼠模型中葡萄糖转运蛋白1过表达可阻止心力衰竭的发生[46]。应用于抗心绞痛、缓解心肌缺血的药物乙莫克舍和哌克昔林可阻碍脂肪酸氧化,同时通过葡萄糖脂肪酸循环促进葡萄糖氧化[47]。
在心脏疾病中针对代谢靶点的治疗方法仍是基础研究及临床应用的重点。目前已有大量通过调节心肌代谢稳态来维护心功能的药物,在心脏疾病的治疗中发挥了重要作用。
心肌细胞通过调节糖脂代谢引起能量变化,以维持细胞稳态。目前研究表明,包括高血压、冠心病、心肌梗死、卒中、慢性心力衰竭等一系列严重的心血管疾病病程发展均伴随着心肌细胞能量代谢方式的转变[3, 14]。
以往研究通常认为,脂肪酸代谢在成年心肌能量中占主导地位,而在某些缺血缺氧状态下,过度的脂肪酸摄取和氧化带来的脂肪酸产物堆积造成脂毒性,诱导细胞凋亡。而在肥大心肌中由于脂肪酸底物不足,以糖代谢为主的功能方式短期内勉强维持心功能,长期却会损伤心肌细胞。乳酸、酮体和支链氨基酸通常不被认为是正常心肌重要的能量来源,大量研究却显示这些代谢底物在心肌细胞能量供应失衡时发挥了生理或病理功能[15]。对于心肌来讲,维持其代谢来源的灵活性和多样性对心肌本身是最有效的保护机制。多种药物已在各类心脏疾病中被证实从代谢稳态角度对心脏疾病的发生发展产生抑制作用。无论是已在临床应用较多的药物,还是针对关键代谢靶点研发的新药,均在心脏疾病的治疗中占有重要地位。
随着代谢组学等检测技术的发展,针对患病心肌代谢产物检测手段和机制探索的研究更加深入。作为我国乃至世界致死率第一的疾病,心血管疾病的防治刻不容缓。在心肌能量代谢方面的深入挖掘,可大大助力对心血管疾病的防治手段的研究。
利益冲突:无
猜你喜欢 糖酵解心肌细胞线粒体 从线粒体动力学探讨中医药治疗心力衰竭相关机制研究世界中医药(2022年18期)2022-11-25线粒体自噬在肠缺血再灌注损伤中的作用及机制研究进展南京医科大学学报(自然科学版)(2022年8期)2022-11-22不同组织来源线粒体提取效率和质量的差异研究重庆理工大学学报(自然科学)(2022年9期)2022-10-26CircRNA-32011调控三氧化二砷诱导的心肌细胞凋亡中国药理学通报(2022年10期)2022-10-14线粒体自噬在纤维化疾病中作用的研究进展中华实用诊断与治疗杂志(2022年1期)2022-08-31circPRKCI靶向miR-217对缺氧/复氧诱导心肌细胞损伤的影响中西医结合心脑血管病杂志(2022年4期)2022-03-11新生SD大鼠心肌细胞原代培养方法的比较牡丹江医学院学报(2022年1期)2022-01-06浅谈400米跑运动员的生化特点和训练读与写·上旬刊(2018年1期)2018-05-06糖酵解中间产物磷酸丙糖鉴定实验课的研究性教学教育教学论坛(2016年51期)2017-03-22“糖酵解”的说课设计教育教学论坛(2016年49期)2017-02-27本文来源:http://www.zhangdahai.com/shiyongfanwen/qitafanwen/2023/0430/591584.html



