【www.zhangdahai.com--其他范文】
白颖慧,许迎浩,吕慎强,李嘉,李惠通,杨泽宇,王林权
(西北农林科技大学资源环境学院,陕西 杨凌 712100)
大气氨(NH3)作为最具流动性的活性氮,极易通过化学反应形成PM2.5和PM10,导致空气质量下降;
大气氨沉降还会加速土壤酸化和水体富营养化,降低物种多样性[1-3]。从1978年到2017年,我国的氨排放量增加了102%,农业活动中过量施肥导致氮肥利用不完全,部分氮以氨的形式损失[4]。草地生态系统具有美化环境、调节小气候、净化空气和水土保持等作用[5]。草地施肥一般采用表面撒施,高温高湿条件更有利于氨气的挥发,其氨损失率与农田相当或更高。有研究表明在草地生态系统中氨挥发损失占氮肥用量的0.04%~60%[6-7]。草地氨挥发受施氮量、灌溉和降雨等影响,同时也受土壤温度、pH、水分、矿质氮含量等影响[8-11]。
植物冠层既是氨源又是氨汇[12],对田间氨挥发具有重要的调节作用。草地成坪后冠层密集,因此,冠层在草地系统氨挥发中的作用不容忽视。植物冠层与大气氨交换量和交换方向主要取决于氨气补偿点及冠层大气氨浓度,环境中的氨气大于氨气补偿点时植物吸收大气氨气,反之则释放氨气[13]。氨气补偿点受质外体NH+4浓度、pH和铵态氮同化的影响,质外体NH+4浓度和pH升高有利于植物释放氨气[14]。植物体铵来源于光呼吸和苯丙氨酸代谢途径[15]。乙醇酸氧化酶(GO)、甘氨酸脱羧酶(GDC)和谷氨酰胺合成酶(GS)等酶活性反映了光呼吸强度,苯丙氨酸解氨酶(PAL)反映苯丙氨酸代谢活性,其与木质素合成有关。另外,植物生殖生长后期营养器官的氮转运也可能与质外体NH+4浓度和冠层氨交换有关[16]。由于C3植物的光呼吸强而C4植物的光呼吸弱[17],因此C3和C4植物的冠层氨交换强度可能存在差异。
目前,关于草地氨挥发有几个科学问题需要明晰:不同植被类型的草地氨挥发通量及其影响因素为何?冠层在草地生态系统中是氨源还是氨汇?C3植物和C4植物之间的冠层交换通量是否存在差异?本试验通过田间原位监测和室内分析相结合的方法,研究3种不同类型草地[暖季型草地狗牙根(C4植物,俗称百慕大)、冷季型草地高羊茅(C3植物)和耐阴冷季型草地麦冬(C3植物)]的土壤氨挥发、冠层氨交换通量、质外体pH、氨气补偿点及其光呼吸和苯丙氨酸代谢途径的相关酶活性,并通过结构方程模型考察冠层氨挥发的生理机制。本研究的主要目的:定量观测3种草地生态系统的周年土壤与冠层氨挥发通量及累积量,以及施肥对其产生的影响;
探索C3和C4草地冠层氨挥发差异及其氨来源。
1.1 试验地概况
试验地位于陕西省杨凌农业高新技术产业示范区西北农林科技大学南校区(34°15"34″N,180°04"18″E),地处关中平原腹地,属于温带季风气候半湿润易旱区,海拔480 m,年降雨量660 mm,年均气温13.0~13.4℃,土壤类型为褐土。土壤基础理化性质见表1。

表1 试验地土壤理化性质Table 1 Soil physicochemical properties of the experimental site
1.2 试验材料与试验设计
供试草地植物品种为麦冬(C3)、高羊茅(C3)和狗牙根(C4),校园内已有种植年份相同的草地,同一草地盖度一致。供试肥料为尿素(N 46.6%)和缓释尿素(河南心连心化肥有限公司生产,属于基质型缓释尿素,通过添加具有较强离子交换性能的功能性吸附材料,控制养分的运移和损失,释放期为50 d,N 45%)。试验设置3个品种,每个品种设3个施肥处理(N0:不施氮处理,SRU:施用30 g·m-2的缓释尿素,U:施用30 g·m-2的尿素),共9个处理,每个处理重复3次,共计27个小区,小区面积为3 m×3 m。试验时间为2021年3月末至11月末,于4月25日将所有氮肥一次性均匀撒施于地表。
1.3 测定项目与方法
氨挥发速率采用泵吸式便携氨气探测仪(Portable ammonia detector,PAD)测定[18]。测定装置由气室(由于草地高度和密度不同,高羊茅和狗牙根采用高20 cm、直径5 cm的PVC管,麦冬采用高25 cm、直径为15 cm的PVC管)、软管和氨气探测仪(博世安电子有限公司)3部分组成。将气室分别罩在土壤或土壤+冠层上,PAD通过泵吸取气室内空气(0.25 L·min-1,由制造商推荐设置),测定其中氨的浓度。各小区随机取点测定,3次重复。施肥后的第一周每天测定,之后间隔5~7 d测定一次。每次测定时间为上午8:00—10:00和下午2:00—4:00,每日氨挥发通量为上、下午的平均值,如有降雨则顺延。
氨挥发通量计算公式:
AE=c×v×14×1 440×104×10-9/(Vm×0.001 962 5)
式中:AE为氨挥发通量,kg·hm-2·d-1(以N计);
c为测定的氨浓度,μmol·mol-1;
v为空气流速,0.25 L·min-1;
14为N的摩尔质量,g·mol-1;
1 440为换算系数,1 d=1 440 min;
104为换算系数,104m2=1 hm2;
10-9为换算系数,109μg=1 kg;
Vm为气体摩尔体积,25℃下为24.5 L·mol-1;
0.001 962 5为气室底面积,m2,测定麦冬时其底面积为0.017 7 m2。
气室内有草的测定值为草地氨通量(土壤+冠层),无草(草与草之间的间隙)的为土壤氨通量。冠层氨通量通过差减法计算,即冠层氨通量=草地氨通量-土壤氨通量
由于冬季氨挥发低于检测限,因此只测定了春、夏和秋季排放量,3季排放量之和为周年排放量。
累积量计算公式:

式中:i为测定次序;
n为总测定次数;
Ai为第i次土壤或冠层氨挥发通量;
Ai+1为第i+1次土壤或冠层氨挥发通量;
N为第i和i+1次之间间隔天数,d。
采集0~5 cm土壤样品测定其硝态氮含量、铵态氮含量、pH和土壤水分。土壤硝态氮和铵态氮采用1 mol·L-1KCl浸提,全自动流动分析仪(AA3,德国)测定,土壤pH采用pH计(水土比为2.5∶1)测定,土壤水分采用烘干法(105℃,12 h)测定,土壤温度采用Fourtec-MicroLite高精度温度记录仪测定。
乙醇酸氧化酶(GO)、甘氨酸脱羧酶(GDC)、谷氨酰胺合成酶(GS)和苯丙氨酸解氨酶(PAL)均采用酶联免疫(Elise)试剂盒(科诺迪生物科技有限公司)测定。质外体pH、NH+4浓度和氨气补偿点采用文献[19]的方法测定并计算。各指标于施肥后每隔7 d测定一次,连续测定3次,之后每月测定一次。
1.4 数据处理
采用Excel处理数据,采用SPSS软件进行单因素方差分析和多重比较(LSD法),采用Amos 22.0(IBM,美国)进行结构方程模型,采用SigmaPlot10.0绘图。
2.1 草地氨挥发
2.1.1 草地土壤和冠层氨挥发
在整个测定周期内,品种和施肥均对土壤氨通量具有显著影响(图1)。两种肥料处理的动态和总量差异不显著,故将二者合并处理,简称N30。N0处理高羊茅、狗牙根和麦冬的土壤氨通量分别为0.174~1.691、0.008~0.107、0.003~0.045 kg·hm-2·d-1,其中,高羊茅的土壤氨通量显著高于狗牙根和麦冬,狗牙根和麦冬之间差异不显著。高羊茅的土壤氨通量为春、秋高,夏季低,狗牙根和麦冬的土壤氨通量整个周年均低而平稳。N30条件下高羊茅、狗牙根和麦冬的土壤氨通量分别为0.074~1.692、0.026~1.587、0.006~0.383 kg·hm-2·d-1;
狗牙根和麦冬的土壤氨通量随施肥显著增加,而高羊茅增加不明显。狗牙根峰值出现在施肥后的第3天,高羊茅在施肥后第6天,而麦冬的峰值出现在夏季(6月)。
在整个测定周期,除了个别时间点外,冠层氨交换以排放为主(图1)。N0情况下,高羊茅的冠层氨通量显著高于狗牙根和麦冬,狗牙根和麦冬之间无显著差异,高羊茅、狗牙根和麦冬的冠层氨通量分别为-0.112~2.547、-0.018~0.155、0.007~0.027 kg·hm-2·d-1。在N30处理下,高羊茅、狗牙根和麦冬的冠层氨通量分别为-0.090~2.171、-0.277~1.518、-0.165~0.266 kg·hm-2·d-1;
狗牙根的氨通量显著增加,而高羊茅和麦冬对氮肥的响应较小。狗牙根和高羊茅冠层氨通量在施肥后的5~6 d达到峰值,而麦冬无明显峰值。
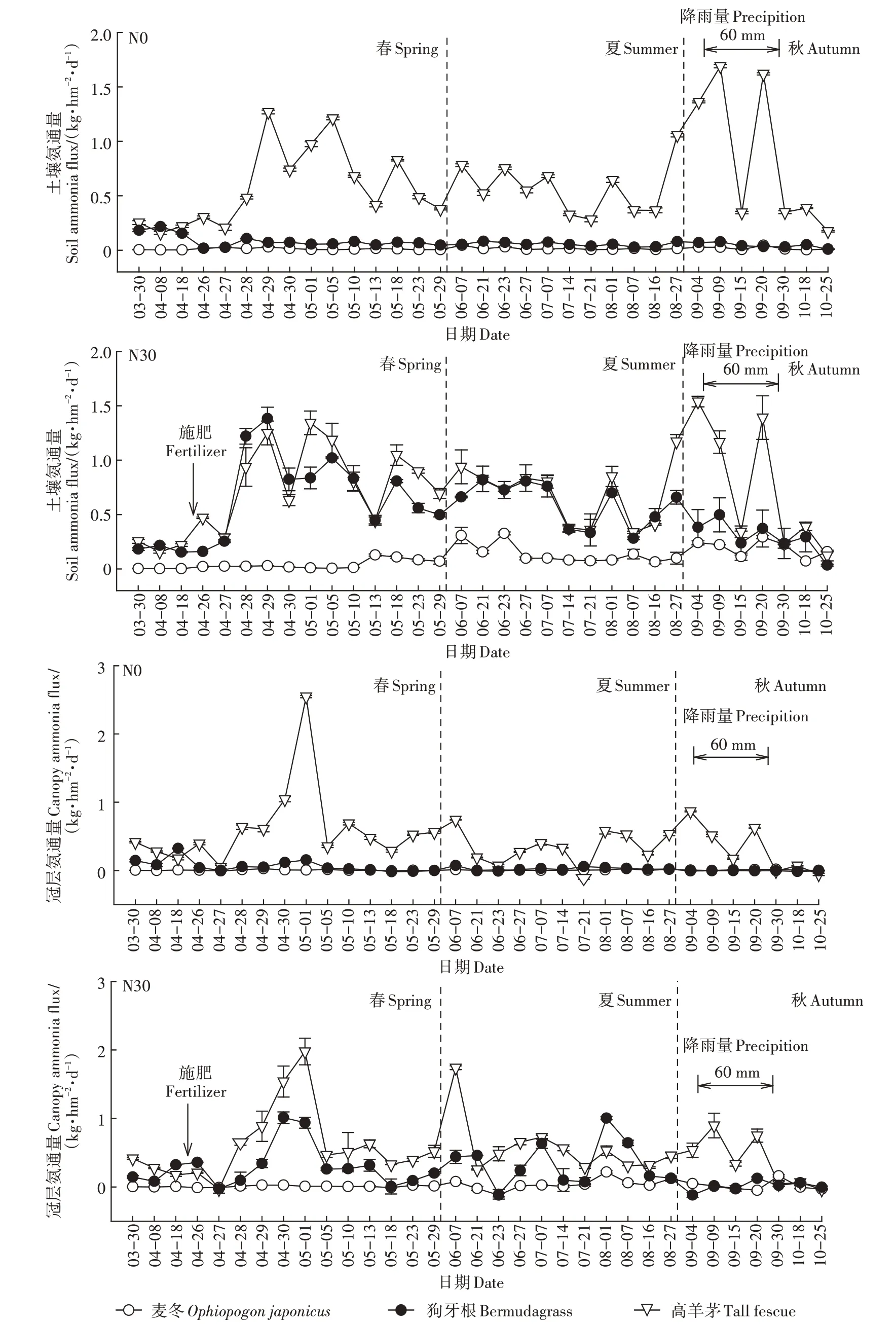
图1 3种草地的土壤和冠层氨通量Figure 1 Soil and canopy ammonia fluxes of three turfgrass species
表2为3种草地的土壤和冠层氨累积挥发量。不同品种之间土壤氨累积挥发量存在显著差异,其顺序为高羊茅(120.9~135.3 kg·hm-2·a-1)>狗牙根(12.5~108.3 kg·hm-2·a-1)>麦冬(2.8~25.3 kg·hm-2·a-1);
与N0(45.4 kg·hm-2·a-1)相比,N30(89.6 kg·hm-2·a-1)显著增加了土壤氨累积挥发量。各品种土壤氨累积挥发量夏季最高,占整个周年的46.2%。

表2 不同季节土壤和冠层氨累积挥发量(kg·hm-2·a-1)Table 2 Soil and canopy ammonia accumulation in different seasons(kg·hm-2·a-1)
不同品种之间冠层氨累积挥发量差异显著,其顺序为高羊茅(79.0~101.6 kg·hm-2·a-1)>狗牙根(7.6~49.3 kg·hm-2·a-1)>麦冬(1.6~8.1 kg·hm-2·a-1);
与N0(29.4 kg·hm-2·a-1)相比,N30(53.0 kg·hm-2·a-1)显著增加了冠层氨累积挥发量。N0处理下,春、夏季氨累积挥发量高,而秋季低;
N30处理下表现为夏季>春季>秋季,冠层氨挥发以夏季为主。冠层氨释放量占草地氨挥发的37.3%(施肥)~39.3%(无肥)。
2.1.2 草地氨累积挥发量及损失率
由表3可知,不同品种之间草地氨累积挥发量差异显著。N0处理的高羊茅、狗牙根和麦冬的氨累积挥发量为199.9、20.1、4.4 kg·hm-2·a-1,N30处理则为236.9、157.6、33.4 kg·hm-2·a-1。与N0相比,N30显著增加了草地氨累积挥发量。夏季、春季和秋季草地氨挥发分别占整个周年的48.6%、29.3%、22.1%。氮肥氨挥发损失率顺序为狗牙根>高羊茅>麦冬,损失率分别为45.8%、12.3%、9.7%,平均损失率为22.6%。

表3 草地的总氨累积挥发量和氮肥的氨损失率Table 3 The total ammonia accumulation loss in field and N fertilizer loss rate
2.2 影响草地土壤氨挥发的因素
3种草地土壤温度均呈先升高后降低的趋势(图2)。狗牙根的冠层密集,有较好的保温效应,因此土温显著高于麦冬和高羊茅,麦冬和高羊茅的土温无差异。狗牙根、高羊茅和麦冬的土壤温度平均值分别为24、21、21℃。如图3所示,土壤pH范围为8.0~8.6,整体规律为先下降后上升最后稳定在8.4~8.5;
与N0处理相比,N30处理土壤pH短期内有小幅度降低。土壤含水量范围为9.6%~31.3%,3—8月随降雨减少缓慢降低,9—10月随降雨量增加而升高,施肥对土壤含水量无显著影响。

图2 草地生长期的土壤温度变化Figure 2 Changes in soil temperature during lawn growth period

图3 草地生长期的土壤pH和土壤含水量变化Figure 3 Changes in soil pH and soil water content during lawn growth period
图4为不同处理下土壤铵态氮和硝态氮的变化规律。N0处理下,高羊茅、狗牙根和麦冬的铵态氮含量范围分别为3.01~6.32、3.09~6.42、2.02~4.90 mg·kg-1,高羊茅和狗牙根的铵态氮含量高于麦冬。N30处理下,高羊茅、狗牙根和麦冬的铵态氮含量范围为2.57~115.08、2.76~82.36、2.33~81.75 mg·kg-1。N0处理下,高羊茅、狗牙根和麦冬的硝态氮含量范围分别为1.96~6.49、0.06~4.13、2.12~5.07 mg·kg-1。N30处理下,高羊茅、狗牙根和麦冬的硝态氮含量范围为1.96~151.67、0.72~88.59、1.39~105.28 mg·kg-1。与N0相比,N30处理显著增加了土壤铵态氮和硝态氮含量。施肥后15 d内土壤铵态氮含量先升高后降低,硝态氮含量从4月28日开始缓慢上升,在5月10日达到峰值后开始下降。

图4 草地生长期土壤矿质氮的变化Figure 4 Changes of soil mineral nitrogen during lawn growth period
2.3 影响冠层氨交换的生理因素
2.3.1 酶活性
从各季节的平均酶活性来看(图5),GO活性随季节变化呈逐步上升趋势。春季各品种GO活性差异不显著,夏季高羊茅和狗牙根显著高于麦冬,秋季高羊茅显著高于狗牙根和麦冬。春季GDC活性显著高于秋季和夏季,秋季高于夏季。春、夏两季高羊茅和狗牙根GDC活性普遍高于麦冬,而秋季高羊茅GDC活性显著高于狗牙根和麦冬。施肥对GO和GDC活性影响不显著。
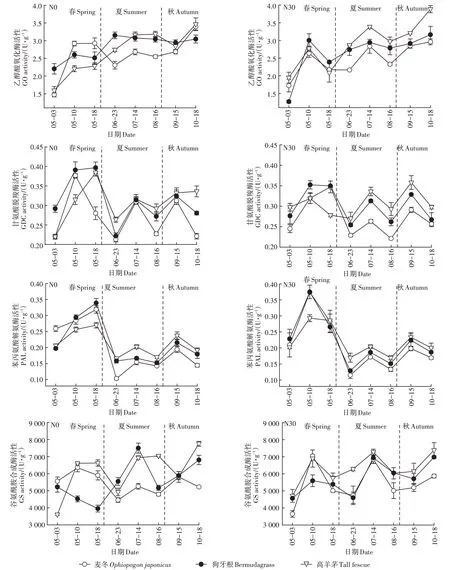
图5 乙醇酸氧化酶(GO)、甘氨酸脱羧酶(GDC)、苯丙氨酸解氨酶(PAL)和谷氨酰胺合成酶(GS)活性的周年变化Figure 5 Annuial activity dynamics of glycolic acid oxidase(GO),glycine decarboxylase(GDC),phenylalanine ammonia lyase(PAL)and glutamine synthase(GS)
春季PAL活性显著高于夏季和秋季,秋季略高于夏季。春季各品种PAL活性差异不显著,夏季高羊茅PAL活性显著高于狗牙根和麦冬,秋季高羊茅和狗牙根PAL活性高于麦冬。GS在整个周年的活性均处于较高水平,夏、秋两季各品种GS活性高于春季。春季各品种GS活性差异不显著,夏季高羊茅和狗牙根GS活性显著高于麦冬,秋季高羊茅GS活性高于狗牙根和麦冬。施肥对PAL和GS活性无显著影响。
2.3.2 质外体pH和氨气补偿点
质外体pH范围为5.9~7.5(图6)。N0处理的高羊茅和麦冬的质外体pH显著高于狗牙根,N30处理的高羊茅的质外体pH显著高于麦冬和狗牙根。与N0相比,N30处理高羊茅和狗牙根的质外体pH增高,而麦冬降低。各处理氨气补偿点呈先升高后降低的变化规律(图6),高羊茅的补偿点高于狗牙根和麦冬。与N0处理相比,N30处理狗牙根和高羊茅的补偿点有所提高,而麦冬补偿点无显著变化。

图6 质外体pH和氨气补偿点Figure 6 Apoplast pH and ammonia compensation point
2.3.3 冠层氨的源解析
图7为C3草地生理指标与冠层氨通量的结构方程模型,高羊茅和麦冬两种C3草坪的分析结果一致,因此合并处理。C3植物的模型(DF=4,P=0.160,CFI=0.989,RMSEA=0.072,模型拟合度良好)显示质外体pH、PAL和GS可以解释43.0%的冠层氨通量变化。其中质外体pH和PAL(标准化回归权重分别为0.596和0.215)为冠层氨交换的主控因素,正向调控冠层氨通量。光呼吸酶GO和GDC通过影响次代谢酶PAL活性间接影响冠层氨通量,GO显著负向调控PAL,而GDC极显著正向调控PAL。PAL、GO和GDC显著正向调控GS。质外体pH正向影响光呼吸酶和GS活性(P<0.05)。

图7 C3植物(高羊茅和麦冬)各生理指标与冠层氨通量的结构方程模型Figure 7 Structural equation model of each physiological index and canopy ammonia flux of C3 grassland grasses(tall fescue and Ophiopogon japonicus)
图8为C4草地生理指标与冠层氨通量的结构方程模型(DF=6,P=0.272,CFI=0.995,RMSEA=0.065,模型拟合度良好)。GDC、PAL和质外体pH及氨气补偿点可以解释74.1%的冠层氨通量变化,其中GDC负向调控冠层氨交换,质外体pH、PAL和氨气补偿点正向调控冠层氨通量。GO通过影响GDC和PAL间接影响冠层氨通量。光呼吸酶GO和GDC极显著正向调控次代谢酶PAL。GO正向影响铵同化酶GS,而PAL负向调控铵同化酶GS。GS和PAL极显著(P<0.001)负向影响氨气补偿点。质外体pH负向影响GO、GS活性,正向影响氨气补偿点和PAL活性(P<0.05)。

图8 C4植物(狗牙根)各生理指标与冠层氨通量的结构方程模型Figure 8 Structural equation model of physiological indexes and canopy ammonia flux of C4 grass(Bermudagrass)
3.1 草地氨挥发
谷佳林等[20]的研究表明施用尿素和不同控释肥(15 g·m-2)后高羊茅草地全年氨累积量为13.59~44.42 kg·hm-2,氮肥损失率为5.23%~17.08%;
边秀举等[21]的研究表明黑麦草施用尿素和不同控释肥(12 g·m-2)后15 d氨挥发损失为0.77%~20.78%;
对匍匐翦股草的研究表明,施用尿素和不同控释肥(14.7 g·m-2)后10 d氨挥发损失率为0~40.0%[7];
狗牙根草地的尿素和不同控释肥(14.7 g·m-2)的氮肥损失率为3.6%~11.7%[22];
也有文献表明0.04%~60%的氮肥以氨挥发的形式损失[6]。本研究结果中麦冬的氨挥发水平与前人研究结果一致,而高羊茅和狗牙根的研究结果(表3)高于前人的研究,其主要原因可能与本试验的施肥量(30 g·m-2)有关。高羊茅草地土壤氨挥发量较高可能与其较高的土壤矿化率有关。高羊茅草地土壤全氮和有机质含量明显高于其他两块草地(表1),可能是土壤有机氮矿化作用高于其他两种草地的原因。图4也表明高羊茅和狗牙根的铵态氮含量高于麦冬(N0)。本研究发现麦冬的土壤和冠层氨挥发均较低,且对肥料的响应也低,因此如果在草皮建设中适当增加麦冬比例,可能有助于氨减排。
本试验施用尿素和缓释尿素处理的草地氨挥发无差异,可能是由于所用的肥料不是包膜控释肥,而是吸附性的缓释肥,其缓释效果在草地上不理想。前人[23-24]的研究表明,尿素与本试验所用的缓释尿素的释放规律基本一致,缓释肥料中添加的基质材料可吸附尿素并抑制溶解。此外本试验施肥采用撒施的方式,肥料暴露在土壤表面,枯草层的脲酶活性比土壤高30倍[25],有利于尿素分解,从而导致缓释肥料没有达到缓释效果。
3.2 草地冠层氨挥发及其贡献
在研究草地氨挥发时一般未区分土壤氨挥发与冠层氨挥发。土壤-大气-冠层之间也存在氨交换,冠层既能释放氨,又能吸收氨。在土壤氨挥发较高时,大气氨浓度升高,当大气氨浓度大于氨气补偿点时就可能产生冠层吸收现象,反之则为氨释放。本研究表明,除个别时期冠层有少量吸收外,大部分时期冠层表现为氨释放。在监测期内冠层氨释放量占整个草地氨挥发的37%~39%,因此仅测定土壤氨挥发会显著低估草地的氨挥发。不同品种草地之间冠层氨挥发存在显著差异,主要是因为其质外体pH、酶活性和氨气补偿点不同。高羊茅的质外体pH、酶活性和氨气补偿点普遍高于狗牙根和麦冬,因此高羊茅的冠层通量大于后者。狗牙根的质外体pH和氨气补偿点对肥料响应高于高羊茅和麦冬,导致施肥对其冠层氨挥发的影响较大。
3.3 冠层氨交换的生理机制与源解析
冠层氨的来源有光呼吸、苯丙氨酸代谢途径和生殖生长期营养器官含氮化合物的转运过程等[15-16]。本研究供试草地植物均没有生殖生长,因此不存在植物体内的氮素转运。光呼吸的甘氨酸脱羧过程释放出一个NH3,PAL在苯丙氨酸生产反式肉桂酸和NH3过程中发挥作用[27-28],这些过程产生的NH3如果未被GS固定将挥发到环境中[15]。从图7和图8的结构方程模型可知C3植物和C4植物的PAL活性均直接与冠层氨通量密切相关,说明两种植物的冠层氨来源均与苯丙氨酸代谢途径有关,这可能与草地乂割导致的茎叶生长过程的木质素合成有关。
C3植物光呼吸的关键酶GO和GDC间接影响冠层氨通量(图7),而C4植物冠层氨通量直接受GDC负向调控(图8)。这说明C3植物和C4植物的氮代谢过程和冠层氨的来源有差异。植物的光呼吸过程发生在叶绿体、线粒体和过氧化物酶中,其中发生在线粒体中的乙醇酸氧化和甘氨酸脱羧通过线粒体电子传递链影响木质素合成酶PAL活性[29-30],同时产生NH3。因此C3植物旺盛的光呼吸作用对冠层氨释放有重要作用。C4植物的光呼吸弱,其产生的氨较少,不会直接影响冠层氨的供应,但是可能通过乙醇酸氧化过程激发GS活性而增加氮同化途径,进而降低质外体pH和氨气补偿点,抑制冠层氨释放。
本研究采用草地原位测定的3种草地土壤肥力具有明显差异,土壤氨挥发也差异显著,如高羊茅草地的土壤氨挥发和冠层氨挥发显著高于其他两种草地,因此冠层氨释放可能受到土壤氨挥发的影响较大。在今后的研究中需要进一步考察同一土壤条件下的C3和C4植物冠层氨交换特征,以消除土壤氨挥发对不同植被冠层氨交换的影响。另外,本试验供试植物种类偏少,今后应该在更多的植物中进行比较研究,并从基因和分子生物学角度探讨C3和C4植物的冠层氨交换规律及生物学机制。
(1)3种植被类型的草地氨挥发通量具有显著的差异,高羊茅、狗牙根和麦冬的氨挥发量分别为199.9~236.9、20.1~157.6、4.4~22.4 kg·hm-2·a-1。
(2)施肥会显著增加草地的氨挥发,其中狗牙根对氮肥的响应显著高于高羊茅和麦冬;
高羊茅、狗牙根和麦冬草地的氮肥氨挥发损失率分别为12.3%、45.8%和9.7%。
(3)冠层氨挥发对草地的氨挥发贡献为37%~39%,冠层氨释放对草地氨挥发的贡献不容忽视。
(4)C3植物和C4植物的冠层氨通量均与苯丙氨酸代谢途径呈正相关,质外体pH对冠层氨通量也有显著影响。C3植物的光呼吸通过苯丙氨酸途径间接刺激冠层氨释放;
而C4植物的光呼吸直接或间接地负向调控冠层氨通量;
氮同化途径通过氨气补偿点和质外体pH间接负向调控冠层氨通量。




