【www.zhangdahai.com--其他范文】
赵权宇
(南京工业大学药学院,江苏 南京 211816)
碳减排是关乎全球长远稳定发展的重大挑战[1]。因此,必须降低大气中的CO2浓度,严格控制全球温度的上升在2℃[2],甚至1.5℃以内[3]。继签订《京都议定书》后,我国政府于2020年9月郑重宣布了“CO2排放力争于2030年前达到峰值,努力争取2060年前实现碳中和”的减碳目标。化学、物理和生物学方法均可进行CO2的封存和利用。在自然界,有6条天然的生物固碳途径[4]。其中,Calvin循环是光驱动的好氧固碳途径,广泛存在于地球上的高等植物和藻类中。英国牛津大学和美国加州大学等单位的科学家在Nature共同撰文评价CO2减排的10条途径,微藻产品是其中之一[5]。为早日达成碳中和目标,研发微藻基生物减排技术大有可为。
微藻通过光合作用固定CO2,在胁迫条件下富集油脂或淀粉等,是高效的光合细胞工厂。近10年来,微藻生物技术在生物燃料等领域的应用受到广泛关注[6]。研究者在藻种选育、光生物反应器设计及大规模培养[7-8]、微藻收获[9]等方面都取得了长足进步。然而,开发微藻基负碳途径还需要克服一些技术瓶颈[10-12]。微藻生物燃料的生产成本仍然较高,用水量大,占用土地广,这极大限制了其产业化步伐。多数微藻不能耐受超过2%的CO2,而工业烟道气中除了10%~25%的CO2,还有NOx和SOx等污染物,这些烟道气组分都会抑制微藻生长[13]。藻种选育是微藻固碳技术路线的起点,藻株强化后有助于达到稳定固碳的目标。为解决微藻产业化层面的问题,可以利用废水资源解决微藻培养中的水需求,可以通过生产高附加值产品提高经济性[14-15]。
除了不断发现新的天然固碳途径[16],合成生物学是强化微藻CO2固定、废水处理和高附加值产品研发,构建微藻基碳中和途径的强大工具。莱茵衣藻[17-18]、三角褐指藻[19-20]和微拟球藻[21-22]等的基因工程改造已经成为可能,然而利用合成生物学手段进行其他真核微藻的途径重构或基因工程改造仍是困难的。很多具有应用潜力的微藻来自野外筛选,还没有进行基因组测序,基因工程改造成功率低。并且,多数环境耐受性并不是简单地由单个基因控制,多靶点的基因改造难度更大。适应性实验室进化(ALE)是合成生物学的重要手段[23-24],已经成功用于细菌、酵母和藻类的菌/藻种选育[25-26]。通过进化工程手段可以提高微藻对环境胁迫的耐受性[27]。其中,开发高附加值产品可提高微藻生物固碳过程的经济性,尚需利用合成生物学手段调控或构建新的代谢途径。微藻是良好的光合固碳底盘,本文将就面向碳中和目标的微藻适应性实验室进化进行综述。
适应进化可以不通过基因工程手段而活化潜在途径、强化特定代谢表型或提高环境适应能力。自然界的生命体在长期演变中为适应环境变化存在自然选择,由此产生适应进化。实验室内,微生物的适应进化实验可以作为研究进化的工具,也可以作为开发工业新菌株的手段[28-29]。
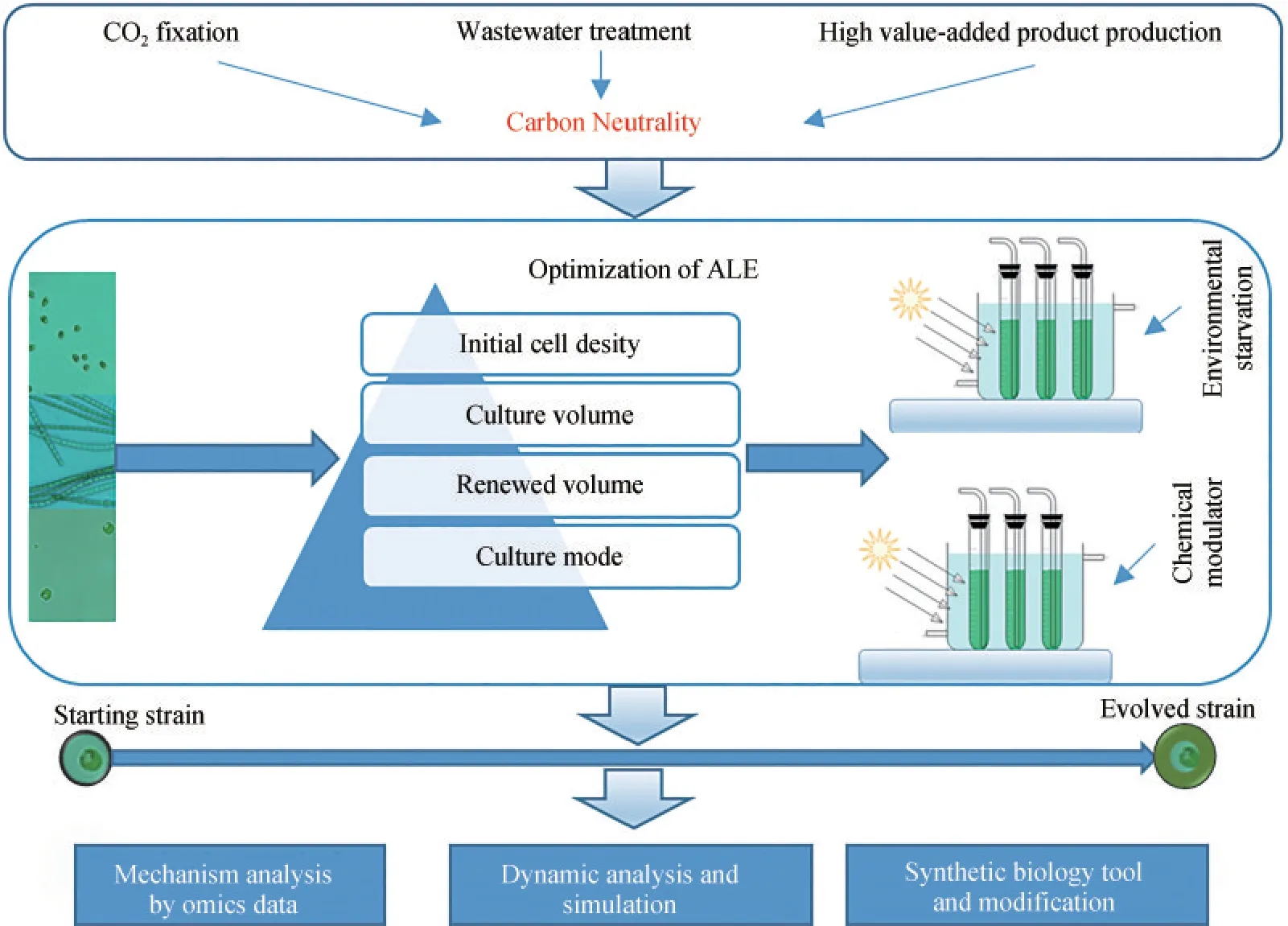
微藻ALE的效率取决于初始藻株的种类、微藻的初始细胞密度和进化策略等(图1)。初始藻株需具备一定的生长、抗逆或高产的基础及潜力。微藻的细胞接种密度、培养体积和每次更新的体积等与细胞增殖密切相关,也影响突变藻株的筛选效率。ALE可为连续培养模式,也可为间歇培养模式。适应进化的常用方法是培养微生物达到一定的指标后进行传代培养。不断重复这个过程直至代谢表型趋于稳定,获得进化藻株。ALE的进化策略包括胁迫策略和非胁迫策略。需要指出的是,胁迫策略是常见的构建抗逆藻株的途径,而非胁迫策略在不抑制生长的条件下,改善生长表型,并提高油脂等的累积[30]。基于化学调节剂的适应性实验室进化(CM-ALE)用于改善隐甲藻的脂质生成[31]。化学调节剂抑制特定细胞内酶的活性,是一种调节碳分配的化学胁迫,显著提高微藻的脂质生产。因此,CM-ALE是构建微藻进化菌株的有效策略。在适应进化中利用富里酸定向调控多不饱和脂肪酸的累积也取得了进展[32]。化学调节剂可在不抑制微藻生长的条件下,定向调控微藻目标产物的合成,是目前的研究热点。改变微藻ALE的藻种选择、基本条件和进化策略,可能会获得不同的进化藻株。微藻的基因突变也是随机的。即使ALE的环境胁迫条件相同,同一个出发藻株经过不同的ALE也会产生不同的进化藻株。即使这些进化藻株的耐受性或进化表型相近,其突变基因也存在差异。

图1 微藻ALE的基本流程Fig.1 Basic procedure of ALE for microalgae
“物竞天择,适者生存”是自然界的普遍规律。适应是能顺应环境胁迫,或同种或异种生物的竞争,最后能生存下来的都属于进化后的“强者”。这也是自然界物种不断演变和极大丰富的原因。在实验室保存的莱茵衣藻也出现了代谢表型的变化[33]。更多的是自然界中响应环境胁迫出现的突变藻株。在意大利Bossoleto二氧化碳泉附近的空气中CO2的浓度可达1080~2160 mg/m3[34],该地的藻株可以耐受3600 mg/m3的CO2。原则上,研究用的藻株都直接或间接来源于自然界,有些在筛选和纯化后进行过诱变等改造。从极端环境等筛选藻株往往具有高耐受性,但是筛选工作量大,费时费力。为了获得可以耐受更高浓度CO2或其他胁迫条件的藻株,ALE是可行的改善微藻代谢表型的技术途径。从适应进化或驯化的时间尺度上,可以将ALE分为短期驯化和长期进化。
2.1 短期驯化
有些研究发现,2~19 d左右的短期驯化就可以获得耐受酸性、高光强、高浓度CO2、高氨氮、高温和高盐度等环境胁迫的藻株。微藻短期驯化的基本条件和时间等总结于表1。相对来说,长期ALE比短期驯化可以获得更稳定和更具耐受性的藻株。经过257 d(1255代)获得的进化莱茵衣藻藻株在0.2 mol/L NaCl的生长高于短期驯化(2 d)的藻株[41]。需要考虑两个问题,短期驯化能否强化耐受表型,以及耐受表型能否稳定遗传。短期驯化能否成功,还要综合考虑藻株和胁迫条件。高盐[42]或烟道气[43]等环境胁迫不能短期获得耐受性,只能采取分段ALE。ALE包括出现不同耐受性突变株,以及高耐受藻株在不同周期中的逐步富集和低耐受株逐步被淘汰的过程(图2)。短期驯化的藻株能否稳定遗传还需要更长期的考察。有必要对短期驯化和长期进化的藻株进行系统分析,探索相关的机制问题。

表1 微藻的短期驯化Tab.1 Short-term acclimation for microalgae
2.2 长期进化
长期进化是获得稳定进化藻株的有效方式。一般来说,微藻在自养条件下生长速度比大肠杆菌或酿酒酵母等微生物在异养条件下慢,在环境胁迫条件下生长周期更长。微藻中的整个适应进化过程可能需要3个月到2年以上的时间。因此,提高适应进化的效率非常重要。微藻长期进化的内容总结于表2。为了提高对环境胁迫的耐受性,适应进化中的初始生物质浓度可能需要0.5~0.8 g/L[57],每个循环(cycle)只能增殖2~3代(generation)。整个适应进化周期增殖代数少,适应进化效率低。一般来说,大肠杆菌或酿酒酵母等微生物的适应进化都在500~2000代。微藻的单一适应进化周期1~21 d不等[32,52]。因为在环境胁迫条件下微藻生长受到抑制,每个周期的最终细胞密度有限。因此,每个周期的增殖世代数可能会很低。为了获得稳定的藻株,总增殖代数是比总周期数更关键的评价指标[27]。提高适应进化效率还有一些新的方法。物理和化学诱变将带来随机的碱基突变,而适应进化过程是对诱变后藻株的再筛选,并修复诱变带来的细胞损伤[58-59]。Kondo教授等[50]结合适应进化和诱变的新方法,虽然接种生物质浓度只有0.02 g/L,84 d后就获得了耐盐藻株。除了对纯微藻的驯化,也有研究采用了对微藻群落的ALE,这样得到的进化藻株更接近实际应用的需要。经过几个月的适应进化,微藻群落可以耐受30% CO2,并在火力发电厂进行了侧线测试[60]。需要注意的是,ALE的时间并非越长越好,进化70 d得到的进化株其生长和油脂含量等指标就略优于进化80 d得到的进化株[61]。为了提高ALE的效率,目前已经建立了微滴培养系统的高通量ALE策略[62-63]。
3.1 CO2固定
微藻培养的CO2来源可以来自空气或者烟道气等工业废气。一般来说,一个藻株有最佳的CO2培养浓度。如果不能耐受高浓度CO2,可以通过ALE强化其对低pH的耐受并改善固碳能力。以10%或
20%CO2为胁迫压力,选择一株野外分离的小球藻为出发藻株,经过30个周期分别获得了两株进化藻株,AE10和AE20[64]。这两株进化藻株在1%~30% CO2条件下均能高效固碳,而且AE10的固碳能力要强于AE20。这也说明,适当的胁迫压力是ALE成功的关键之一,并非胁迫压力越大获得的进化藻株越好。烟道气中的NOx和SOx在微藻培养基中累积,会快速降低培养基pH,同时也会存在盐离子毒性。选择经过一次ALE的进化藻株AE10作为新的出发藻株,以模拟烟道气作为胁迫压力,分5段递增模拟烟道气浓度进行适应进化后,获得了新的进化藻株Cv(二次进化藻株),可以耐受10%CO2、200 mL/m3NOx和100 mL/m3SOx[43],为微藻直接利用烟道气进行固碳提供了可能。需要指出的是,工业烟道气中还含有其他痕量污染物,对微藻也存在抑制作用[13]。研究发现,进化藻株AE10是通过10%CO2作为胁迫压力经过ALE获得的,但是AE10具有耐受高光强[65]和20 g/L NaCl[42]的多种抗逆性能,再经过两段适应进化,获得了可以耐受30 g/L NaCl的进化藻株S20[36]。从一个淡水藻株,到一个可以在海水中生长并快速固碳的藻株,ALE发挥了重要作用。
3.2 废水处理
微藻培养需要大量的水,而工农业废水和城市污水中含有一定浓度的氮、磷,是微藻培养必需的营养元素。利用废水培养微藻可以构建绿色生态循环,也有助于实现双碳目标。高浓度苯酚会抑制微藻生长,出发藻株L3经过90 d的ALE,进化后的小球藻藻株L5可以去除500~700 mg/L的苯酚[57]。ALE也成功用于强化球等鞭金藻(I.galbanaParke)对苯酚的耐受性[51]。先在100 mg/L苯酚的胁迫下进化90 d,得到进化藻株PAS-1,再在200 mg/L苯酚的胁迫下进化90 d,得到PAS-2。PAS-1和PAS-2可以分别耐受200 mg/L和300 mg/L的苯酚。并且,这两个进化藻株均可在5 d完全去除200 mg/L的苯酚。从垃圾渗滤液中分离得到藻菌菌群,经过2年的ALE,群落中小球藻的生长速率提高了两倍,硝态氮和氨氮均可去除[53]。废水和污水中往往含有细菌等微生物,开放式培养更是无法做到完全无菌,菌群的适应进化可以构建更具鲁棒性的培养系统[66-67]。废水中一定浓度的有毒有害物质,对微藻构成胁迫。有可能通过ALE强化耐受性并高效去除这些污染物。废水的种类众多,污染物的种类和浓度也千差万别,ALE用于强化微藻废水处理藻株的研究还有必要进一步深入。
3.3 表型强化
表型强化包括强化生长表型和强化代谢表型。野生型大肠杆菌的生物质合成只能达到理论计算值的40%~60%。刚刚构建的单基因敲除突变体甚至无法正常生长。这些突变体经过适应进化,生物质合成或代谢产物生产等均会显著增加,与基因组尺度代谢网络模型的预测结果接近[68]。微藻固碳之后,细胞内将进行碳分配,调控获得更多的油脂或糖。理论上,抑制糖合成将提高油脂的累积。但是,经诱变获得的两株莱茵衣藻糖合成缺陷突变株的油脂含量并未显著提高[30]。在最佳生长条件下,以该两株藻作为出发藻株,经过84 d的ALE,进化后藻株的生长均得到强化。并且,其中一进化藻株在缺氮条件下的油脂含量与进化前相比提升了将近1倍。ALE除了强化了生长,对油脂[69]和色素的合成[46-47]也有促进作用。环境胁迫条件将在微藻胞内产生活性氧自由基,并调控碳分配向DHA等多不饱和脂肪酸[54-56]。微藻油脂等高附加值产品的累积往往通过培养过程的环境胁迫来获得,但是环境胁迫会限制微藻生长。如果采取两段法,第一段生长,第二段油脂或糖等累积,两段法的总体时间过长会降低产率[65]。如果通过ALE获得进化藻株,可以既不抑制微藻生长,又高产高附加值产品,将进一步提高微藻固碳技术途径的可行性。
4.1 机制分析
进化前后的藻株在生理生化[70]、基因水平[71]、转录水平[72-73]、代谢物浓度[45]和表观遗传[58]等方面均存在差异。出发藻株和进化藻株的比较组学分析有利于理解进化藻株对环境胁迫的响应与耐受机制[74]。微生物的适应进化多数是点突变,进化前后的菌株只在1~80 bp碱基上存在差异,只有在个别情况下是大片段基因变化造成的表型差异[75]。而且,可能有多个基因型具有相似的表型。酿酒酵母耐受盐酸和乳酸的研究也表明有多个途径都可以实现耐酸表型[76]。藻类的基因组比微生物大得多,基因注释也不完善。适应进化前后比较基因组学研究将从根本上揭示适应进化的分子机制。此外,需要从热力学等角度认识适应进化过程。黄和等[54-56]构建了多个裂殖壶菌的进化株。综合分析这些裂殖壶菌适应进化藻株的发酵过程。结果表明,DHA发酵过程的有效能效率与生物量产率存在线性关系(R2=0.8044),说明适应进化可以通过强化生长表型提高DHA有效能效率,为进一步通过适应进化提高DHA发酵性能提供参考[77]。
转录组学可以在全基因组尺度比较出发藻株和进化藻株的实时差异。光合作用对微藻生长具有重要影响。一般的环境胁迫可能会对光合系统活性造成损伤,使得微藻的生长受到抑制。对比耐受苯酚进化藻株L5和出发藻株L3,比较转录组结果表明适应进化确实强化了代谢网络中多数基因[70]。但是苯酚的存在仍会下调光合作用、氧化磷酸化和部分氨基酸代谢相关基因的表达。耐受苯酚的机制来源于光合作用、抗氧化酶和色素合成等相关基因的上调[70,78]。对可耐受30 g/L的盐的适应进化藻株S30和AE10进行无参转录组分析[42]。结果表明,高盐度严重影响光合作用、氧化磷酸化、脂肪酸合成和酪氨酸合成代谢,相关基因显著下调。抗氧化酶、CO2固定、部分氨基酸代谢、中心碳代谢和ABC传递系统显著上调。值得注意的是,除了C3固碳途径,C4固碳途径和景天酸代谢途径的基因都显著上调。这说明相关途径与耐盐相关。耐受模拟烟道气的进化藻株Cv在模拟烟道气和10%CO2中的比较转录组结果中显著变化的基因仅有495个,集中在光合作用、氧化磷酸化和氨基酸代谢等代谢途径中[43]。进化藻株Cv在模拟烟道气中通过上调光合作用、氧化磷酸化、氮还原和硫代谢中cysA基因,快速利用氮和硫,防止溶解在培养基中的NOx和SOx降低pH,保护微藻光合系统。耐受苯酚[70]、盐[42]和模拟烟道气[43]的适应进化转录组学机制研究结果表明,耐受性均涉及光合系统、抗氧化等多个代谢途径中多个基因的显著变化,进行多基因的合成生物学改造存在困难。经过650天ALE得到可耐受高浓度葡萄糖的寇氏隐甲藻(C.cohnii)进化藻株[52]。代谢组学分析结果表明,甘油、谷氨酸、丙二酸和琥珀酸是中枢代谢物。这些组学分析获得了微藻的一些代谢调控规律和耐受机制,比较基因组学的研究可以获得更多基因或元件信息,为合成生物学改造提供借鉴。
4.2 进化工程的动态研究
为深入理解适应进化的机制,需要考察适应进化的动态过程。在ALE过程中,随着周期的增加,产生不同的突变株,而且突变株种群也在不断变化(图2)。在环境胁迫条件下,能够适应的突变将保存下来,并在种群内部占据主体,最后可能得到单一突变体,或者得到几个突变体的组合。如果能快速鉴别和筛选这些高耐受突变体,将显著提高ALE效率。在提高耐受性的ALE中,环境胁迫压力是主要的筛选条件,如果是面向提高生长和代谢物高产表型的ALE,则需要研发有针对性的检测指标或生物传感器。
为了更好地理解ALE的动态过程,已经开发一个ALE的优化程序,ALEsim[79],为进一步设计和优化ALE提供了工具。ALEdb 1.0则是一个微生物ALE的网上数据库[80],该数据库包含11000个突变体的信息。通过以往这些ALE的成功实验结果,可以更精准地设计微藻ALE过程。需要注意的是,部分文献中关于ALE过程的描述不够细致,或者根本没提供ALE过程的详细数据,很难进行系统比较。这也要求在微藻的ALE实验中采集足够的信息,为今后的分析打下基础。更需要从酵母[81-83]或细菌的ALE中借鉴经验,完善微藻ALE的设计[24,83]。
4.3 合成生物学在ALE中的应用
近年来,合成生物学在基因组编辑[84-86]、代谢调控[87-88]等方面都有了突破性进展。合成生物学和进化工程结合[89],将提高微藻ALE的效率,构建更有抗逆性和固碳能力的藻株。
ALE为合成生物学提供新的抗逆基因或元件。ALE后获得的进化藻株中存在一定的基因突变,通过比较基因组学等手段可以获得潜在的抗逆基因或代谢工程改造的候选基因。相关元件的挖掘将为进一步的合成生物学改造提供参考[90]。生物元件是合成生物学的基本要素之一[91]。目前,利用比较基因组学等手段,获得了微藻耐受低温等的基因和元件信息。在极地雪藻(Chlamydomonas nivalis)从22℃转到4℃培养时,12个转录因子和3个RNA键合蛋白出现显著变化,积极调控微藻进行响应[92]。通过基因组、转录组和蛋白质组等多组学数据全面分析了可在接近0℃生长的小球藻的基因、转录和蛋白等水平的差异,进一步的发掘将会获得新颖的生物元件[93]。通过合成生物学手段强化光合系统效率,对微藻的藻种改造至关重要[94]。在蓝藻中获得的相关元件也可为真核微藻借鉴。
从进化藻株筛选的候选抗逆基因等还需要进行验证。已从自然进化的藻株中发现了基因变异[71,95]。在低浓度CO2的环境中,微囊藻(Microcystis)的无机碳利用基因以bicA+sbtA为主。当CO2浓度增加,则以bicA为主[95]。在模式微生物中发现候选抗逆基因,通过合成生物学手段改造微生物,再通过ALE强化表型是比较成熟的菌种改造路线。比如,柠檬烯会抑制解脂耶氏酵母生长,造成柠檬酸产量低。通过转录组学发现82个基因上调响应柠檬烯胁迫。从中选择8个候选基因进行改造,确实获得了高产柠檬烯的酵母菌株[96]。微藻的转录组等组学信息也揭示了一些显著变化基因,可以采取类似策略进行改造。但是真核微藻的合成生物学改造在模式微藻中成功率较高[97]。对非模式真核微藻很难通过反向代谢工程手段进行验证。主要原因是受限于真核微藻基因组信息和基因操作工具的缺乏、低效。比如微藻同源重组效率低[98-99],而非模式微藻的基因组编辑工具尚需完善[100]。微藻的代谢调控机制非常复杂,往往是多个基因协同作用,而多基因的基因操作就更为困难。此外,真核微藻中光合作用也非常重要,但是酿酒酵母和大肠杆菌均无光合系统,蓝藻等原核生物的基因操作较为成熟,可以考虑在蓝藻中验证。利用合成生物学手段进行多位点编辑,可以获得具有遗传多样性的进化菌株,也是提高ALE效率的潜在方法[24]。基因组多位点编辑在酵母中应用较广[101],但是对于微藻还是困难的。随着技术的发展,非模式真核微藻的基因组编辑工具将更高效,其基因组多位点编辑也会成为可能。
微藻通过光合作用进行固碳,在自然界中为碳循环贡献着力量。面向“双碳”重大战略需求,需要发挥微藻的优势,提出可行的碳中和路线。根据这些技术路线的需要,ALE强化藻种代谢表型和耐受性,并提高这些技术路线的技术可行性和经济可行性。目前,微藻的ALE还存在效率低、机制分析不明确、合成生物学应用少等问题。
(1)进一步提高微藻的固碳效率,服务碳中和目标。微藻固碳的利用率和对不同工况的适应性还有待提高。部分进入培养系统的CO2停留时间短,造成对CO2的利用率低。从光生物反应器的结构设计等方面可以提高碳的利用率,对藻种来说,微藻快速生长就可以高效固碳。还需要提高微藻对不同工况的适应性,特别是室外培养中温度和光强等周期性变化。在培养体系中加入CO2吸收剂也是增加CO2溶解度和固碳效率的方法,有些吸收剂对微藻也会产生抑制,需要通过ALE强化对这些吸收剂的耐受性。
(2)提高微藻废水处理效率,满足微藻对水资源需要同时有效固碳。微藻废水处理中除了氨氮和盐胁迫,不可避免地还要受其他化合物和微生物的影响。对于含菌系统如果提高微藻的生长,发挥藻和菌各自的作用,也可以通过ALE构建更稳定的体系,进而提高微藻废水处理效率。
(3)综合开发微藻高附加值产品,提高碳中和路线的经济性。ALE除了可以提高微藻的生长,并高效固碳,还提高了油脂或者色素的产率。为了进一步提高微藻碳中和技术路线的经济性,还需要筛选新的微藻高附加值产品。可以通过加入化学诱导剂的方法提高这些产品的产率,相关的策略还有待进一步完善。
(4)探索微藻ALE的分子机制和代谢调控规律。微藻进化藻株的机制分析从根本上来说,还要通过比较基因组和表观组学的方法进行分析。目前相关研究还不充分,因为微藻的基因组测序数量比微生物少得多,基因注释还不完善。转录组学、代谢组学和蛋白质组学等更多的是从代谢调控角度阐明相关机制。转录组及代谢组的研究一般仅停留在代谢网络层面,对信号传导途径的研究不多。抗逆性与信号传导途径息息相关。构建多组学的系统生物学分析,将有助于明确对微藻代谢和抗逆机制的认识,并为进一步的合成生物学研究服务。
(5)提高微藻ALE的效率。随着微藻ALE研究的深入,特别是可以借鉴其他微生物ALE的成果,从藻株筛选、ALE条件和进化策略等角度合理设计,努力使微藻尽快达到ALE的终点,获得进化藻株。搜集ALE中的动态信息并构建模型,获得ALE的进化规律。可以和诱变等方法结合,构建更多的突变体,并高效筛选满足耐受性的藻株。加入适当浓度的化学调节剂,可以在不抑制微藻生长的前提下,调控胞内代谢产物累积,也有利于提高微藻ALE效率。
(6)发挥合成生物学的优势,构建非天然固碳途径。以微藻作为可光合作用的底盘,通过进化工程强化藻株的耐受性,并获得更多的进化藻株。通过比较基因组学分析等,发掘固碳和抗逆元件,可以在其他生物中进行异源表达,并合成新的非天然代谢途径,为实现碳中和目标贡献力量。随着基因编辑技术的发展,真核微藻的基因工程改造效率将不断提高。研发新模式微藻的底盘细胞,并通过合成物学手段构建固碳和高附加值产品生物合成新途径,将助力微藻合成生物学技术实现碳中和目标。
猜你喜欢微藻耐受性表型代食品运动中微藻的科研与生产古今农业(2022年1期)2022-05-054个地被菊新品系对湿热胁迫的耐受性研究园林科技(2020年2期)2020-01-18建兰、寒兰花表型分析现代园艺(2017年21期)2018-01-03絮凝法采收生物燃料微藻的研究进展环境保护与循环经济(2017年3期)2017-09-26巴氏醋杆菌核酸修复酶UvrA对大肠杆菌耐受性的影响天津科技大学学报(2016年1期)2016-02-28GABABR2基因遗传变异与肥胖及代谢相关表型的关系中国康复理论与实践(2015年10期)2015-12-24慢性乙型肝炎患者HBV基因表型与血清学测定的临床意义医学研究杂志(2015年5期)2015-06-10miR-29b通过靶向PI3K/Akt信号通路降低胃癌细胞对顺铂的耐受性中国医学科学院学报(2015年5期)2015-03-01阿米替林治疗脑卒中后中枢痛的疗效和耐受性分析中国医药科学(2015年15期)2015-02-2772例老年急性白血病免疫表型分析现代检验医学杂志(2015年5期)2015-02-06本文来源:http://www.zhangdahai.com/shiyongfanwen/qitafanwen/2023/0912/653116.html



