【www.zhangdahai.com--其他范文】
杨金宏,谢满超,文欣茹,陈蕊茹,孔卫青
1. 安康学院现代农业与生物科技学院/陕西省茶叶省市共建重点实验室,陕西 安康 725000;
2. 安康学院陕西省蚕桑重点实验室,陕西 安康 725000
茶网蝽(Stephanitis chinensis)为半翅目(Hemiptera)异翅亚目(Heteroptera)网蝽科(Tingidae)昆虫,不完全变态,以我国西南地区的油茶和茶树等经济作物为主要寄主。茶网蝽若虫和成虫以刺吸式口器从茶树树冠中、下层成熟叶片背面吸食汁液,其排泄物在叶片上形成黑色黏质物[1],受害茶树树势衰弱,生长缓慢,发芽率低且芽包小,严重影响茶树生长和茶叶产量[2]。
茶网蝽在陕南地区的茶叶产区一年发生2~3代,繁殖力强,群体数量庞大,种群扩散能力强,新旧茶园均受危害。2010年陕西省汉中市首次报道茶园发生茶网蝽危害[3],2012年扩散至陕西省安康市,2017年蔓延至汉江流域和嘉陵江流域[4]。茶网蝽成虫体长 3.1~4.2 mm,体小扁平,无趋性,初孵若虫肉眼无法识别,亟待分子生物学鉴定方法[5]。
线粒体基因组(Mitochondrial genome,mitogenome)不易发生重组、倒位、易位等突变,结构保守。线粒体基因进化速率快,为核基因的 2~6倍,被广泛应用于种群多样性鉴定、物种系统发育、物种地理学谱系和系统发生学等方面的研究[6-7]。全世界已知网蝽科昆虫有260属2 124种,但目前NCBI公布的线粒体基因组仅27个(截至2022年3月31号),网蝽科昆虫的线粒体基因组大小差异较大,主要是由于控制区(Control region)数量、长度不等的重复序列造成[8]。本研究利用Illumina和Sanger测序,获得了陕西安康茶区茶网蝽的线粒体基因组全序列及特征,并进行了网蝽科的系统发育分析,旨在为进一步对茶网蝽扩散传播途径、防治及异翅亚目的分类研究提供依据。
1.1 试验材料
茶网蝽成虫于2021年5月17日采集自陕西省安康市紫阳县和平茶厂茶园,采集的样本存放在无水乙醇中,置于-20℃冰箱保存备用。
1.2 基因组DNA提取与高通量测序
取乙醇浸泡的茶网蝽单个个体,ddH2O冲洗2遍,然后使用快速DNA提取检测试剂盒[KG203,天根生化科技(北京)有限公司]按照说明书提取基因组 DNA,最后用100 μL ddH2O洗脱。使用 Multiskan FC微量酶标仪(美国Thermo Fisher Scientific公司)对提取的基因组DNA进行检测。选取部分检测合格的基因组DNA委托生工生物工程(上海)股份有限公司进行建库和Illumina Hiseq 2500平台双向高通量测序,测序长度为150 bp,目标数据量4 Gb。
1.3 序列拼接组装与控制区的PCR扩增
对Illumina高通量测序数据进行质控,去除接头和带 N的序列,滑窗法去除得分低于20和长度小于 35 bp的序列。最后数据利用GetOrganelle[9](K=31、55、85)和NOVOPlasty[10](K=31)软件进行组装,组装结果使用Bandage[11]进行可视化分析,手动去除可信度低和不确定的部分。在获得的序列缺口处设计引物rrnS-F 和 trnM-R(表1),使用 Gloria Nova HS 2×高保真Taq DNA聚合酶(RK20715,武汉爱博泰克生物科技有限公司)进行 PCR扩增,PCR反应程序:94℃预变性 2 min;
94℃变性 30 s,55℃退火 30 s,72℃延伸 5 min,30个循环;
72℃延伸10 min。使用1%琼脂糖凝胶电泳检测PCR产物,之后对产物进行 Sanger双向测序,并根据获得序列设计测序引物(表1)进行Gap补齐。

表1 引物序列Table 1 Primers sequence
1.4 基因组注释和分析
使用在线软件MITOS2[12](http://mitos2.bioinf.uni-leipzig.de/index.py)对茶网蝽线粒体基因组进行注释,遗传密码选择5(Invertebrate)。同时利用blastn和blastp比对网蝽科其他昆虫的线粒体基因组数据确定蛋白质编码基因(Protein-coding genes,PCGs)、tRNA 和 rRNA的边界位置。最后的注释文件提交至NCBI数据库,并利用在线 OGDRAW 软件[13](https://chlorobox.mpimp-golm.mpg.de/OGDr aw.html)绘制结构图。tRNA的二级结构预测使用在线软件tRNAscan-SE[14](http://lowelab.ucsc.edu/tRNAscan-SE),未成功预测的二级结构进行手工绘制。
使用在线工具 Tandem Repeats Finder[15](https://tandem.bu.edu/trf/trf.html)查找控制区中的串联重复序列,使用MISA[16]检测其中的简单重复序列(Simple sequence repeats,SSRs),参数设置为1~6碱基单元的最少重复数分别为10、5、4、3、3、3个。茎环结构预测使用在线工具MFold[17](http://www.unafold.org/mfold/applications/rna-folding-form.php)。
使用 Mega 11软件[18]统计基因组及各区域 4种碱基的组成和蛋白质编码基因的密码子情况,并通过公式AT skew=(A-T)/(A+T)和 GC skew=(G-C)/(G+C)[19]计算核苷酸组成的偏向性。
1.5 系统发育分析
从 NCBI数据库下载异翅亚目昆虫线粒体基因组,分别提取13个PCGs序列,然后利用 macse_v2.05软件[20]基于编码框架模式对序列进行比对,并把对齐的DNA合并成一个序列。利用 raxmlHPC[21]构建极大似然(Maximum likelihood,ML)发育树,GTRGAMMA模式,抽样 1 000次计算bootstrap值,展示高于50%的位点。
2.1 茶网蝽线粒体基因组的测序与组装
Illumina测序数据经质控,获得2 025 131条序列,GetOrganelle和NOVOPlasty组装软件获得的线粒体基因组序列完全一致的部分长度为14 561 bp,平均深度为415.04倍。控制区序列的组装得到多种结果,可能存在重复结构。进一步利用引物rrnS-F和trnM-R进行PCR扩增,获得1条4 000 bp左右的条带(图1)。使用设计的多条测序引物对控制区进行Sanger测序,拼接测序结果,获得序列长度为4 040 bp,与Illumina测序组装数据拼接成环,获得茶网蝽线粒体基因组全长为 18 085 bp(GenBank登录号:OM397399)。

图1 rrnS-F和trnM-R引物的PCR扩增Fig. 1 The PCR amplification of rrnS-F and trnM-R
2.2 茶网蝽线粒体基因组注释与结构
分析发现,茶网蝽线粒体基因组为典型的双链闭合环状结构,包含37个基因,分别为13个PCGs、22个tRNA基因和2个rRNA基因(图2,表2)。基因组控制区为3 678 bp,其后为trnI(GAU)、trnQ(UUG)、trnM(CAU)和nad2基因,与昆虫线粒体祖先基因顺序(Ancestral gene order)相同[22]。

图2 茶网蝽线粒体基因组图谱Fig. 2 Map of S. chinensis mitogenome
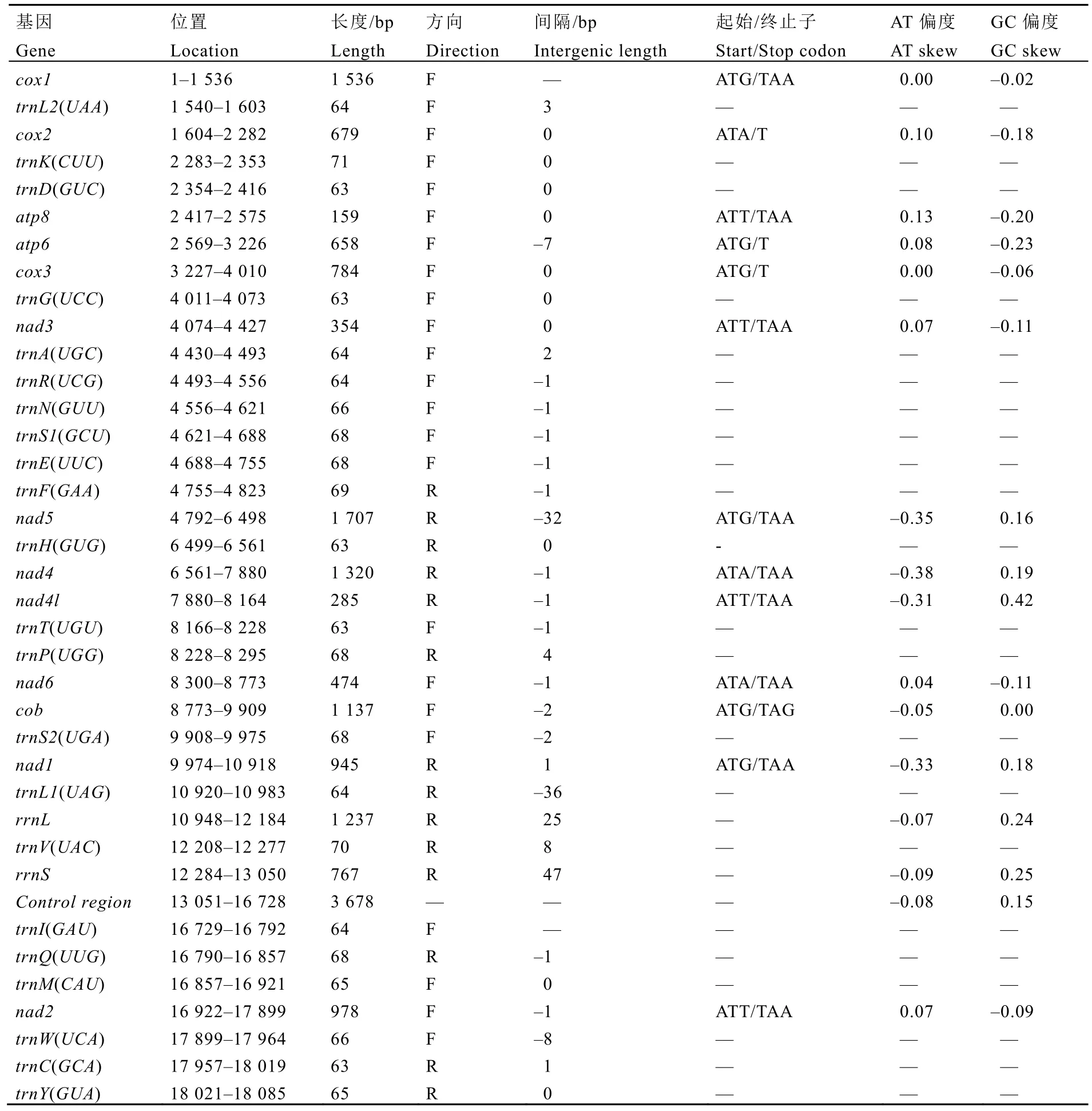
表2 茶网蝽线粒体基因组结构Table 2 The mitogenome organization of S. chinensis
茶网蝽的线粒体基因组的37个基因共存在18处基因重叠,最长的两处位于nad1和trnL1(UAG)、trnF(GAA)和nad5之间,重叠长度分别为-36 bp和-32 bp,其他位置的重叠长度均在-8 bp以内。基因间隔现象少于基因重叠,37个基因仅存在8处间隔,其中最长两处位于trnV(UAC)和rrnS、trnL1(UAG)和rrnL之间,长度分别为47 bp和25 bp,其他间隔最长8 bp,另有无间隔和重叠的区域10处(表2)。
2.3 茶网蝽线粒体基因组核苷酸组成
茶网蝽线粒体基因组的 AT含量为78.09%,具有明显的AT偏好性(表3)。rRNA基因的AT含量最高,为80.84%,AT富集区为79.99%。PCGs的AT含量最低,为76.76%,其中atp8基因的AT含量最高,达84.28%,其次是nad2和nad4l,分别为 82.21%和80.70%;
AT含量最低的是cox1和cob,分别为69.08%和72.21%。核苷酸组成在线粒体基因组不同链间也不对称,多数编码基因所在的J链(Majority strand)的PCGs多为AT偏移和CG偏移,而N链(Minority strand)的PCGs和rRNA基因,为TA偏移和GC偏移(表2)。

表3 茶网蝽线粒体基因组核苷酸组成Table 3 Nucleotide composition of the mitogenome of S. chinensis
2.4 蛋白质编码基因
茶网蝽线粒体基因组的13个PCGs的长度在 159~1 536 bp,其中nad1、nad4、nad4l和nad5位于基因组N链,其他9个均位于J链(表2)。13个基因中,cox1、cox3、atp6、nad5、cob和nad1以ATG起始,atp8、nad3、nad4l和nad2以 ATT起始,cox2、nad4和nad6以ATA起始;
以TAA为终止密码子的基因最多,共有9个,cox2、atp6和cox3以不完全的T终止,cob以TAG终止。同时,PCGs之间存在相互重叠的现象,atp8和atp6基因间重叠了7 bp,nad4和nad4l基因、nad6和cob基因分别重叠了1 bp和2 bp。
进一步分析发现,茶网蝽线粒体PCGs使用最多的 5个密码子分别是 UUA、AUU、UUU、AUA和 AAU,均由 A和 T组成(表4)。编码蛋白的氨基酸组成及含量也有较大差异,使用频率最高的为亮氨酸(Leu),占比约14.00%,其次为异亮氨酸(Ile)、苯丙氨酸(Phe)、丝氨酸(Ser)、甲硫氨酸(Met)、天冬酰胺(Asn)。其他氨基酸占比均低于5%,其中精氨酸(Arg)、半胱氨酸(Cys)、谷氨酰胺(Gln)、组氨酸(His)、天冬氨酸(Asp)的占比低于2%。

表4 茶网蝽线粒体基因组蛋白编码基因相对同义密码子使用度Table 4 Relative synonymous codon usage (RSCU) in the mitogenome of S. chinensis
2.5 tRNA和rRNA基因
茶网蝽线粒体基因组包含22个tRNA基因(图3),长度在63~70 bp。8个tRNA基因(trnC,trnF,trnH,trnL1,trnP,trnQ,trnV,trnY)位于基因组N链,其余14个位于J链(表2)。预测 tRNA基因的二级结构发现,trnS1(GCU)缺少DHU臂,其他tRNA均能形成典型的三叶草结构(图3),该现象存在于多数后生动物中[23-24]。同时,部分tRNA的二级结构还存在G-U和U-U等非经典配对,该现象在其他昆虫类群 tRNA二级结构中也较为常见[25-26]。

图3 茶网蝽线粒体基因组tRNA预测二级结构Fig. 3 Predicted secondary structures of tRNA genes in mitogenome of S. chinensis
茶网蝽线粒体基因组含有 2个 rRNA基因,rrnL位于trnL1(UAG)和trnV(UAC)之间,rrnS位于trnV(UAC)和控制区之间,长度分别为1 237 bp和767 bp,AT含量分别为80.22%和81.88%,具有明显的AT偏向性。
2.6 控制区分析
茶网蝽线粒体基因组的控制区位于rrnS和trnI(GAU)基因之间,长度为 3 678 bp,根据结构将其分为两部分(图4)。第一部分位于序列的前端(1~2 447 bp,front-end region,FER),存在以下结构:(1)3组非串联重复序列。第一组长度为1 540 bp(R1),包含2个498 bp的完整重复单元和 2个该单元的部分序列(477 bp和 63 bp);
第二组长度为 154 bp(R2),包含2个77 bp的完整重复单元;
第三组长度为582 bp(R3),由3个194 bp的完整重复单元组成。(2)4处(TTAG)n,其中(TTAG)7和(TTAG)9分别连接非串联重复 R2的 2个单元,另 2处均为(TTAG)5,位于 R2重复的2个单元中。第二部分位于序列的后端(2 448~3 678 bp,back-end region,BER),为串联重复区(R4),包含5个长度为237 bp的完整重复单元和1个该单元的部分序列。同时序列存在5种茎环结构,其中2种5处为单一环结构,另3种11处为多环结构。

图4 茶网蝽线粒体基因组控制区结构Fig. 4 Structure of mitogenome control region of S. chinensis
进一步分析该区域的AT含量为79.99%,高于Pseudacysta perseae、Agramma hupehanum和杨柳网蝽(Metasalis populi)该区的AT含量,低于其他网蝽科昆虫。FER和BER的AT含量分别为79.85%和80.26%(表3),两段区域对不同核苷酸的使用偏好也有所不同,FER的 AT偏度为-0.12,发生 TA偏移,BER的AT使用量相当,有GC偏移倾向。
2.7 系统发育分析
以双线铜翅沫蝉(Aeneolamia contigua)为外群,构建了基于异翅亚目41种昆虫的线粒体PCGs的系统发育树(图5)。结果表明,茶网蝽与直脊冠网蝽(Stephanitis mendica)的亲缘关系最近,其次为杨柳网蝽和Pseudacysta perseae。网蝽科聚为一簇,位于发育树的根部,是臭虫次目(Cimicomorpha)盲蝽科(Miridae)、花蝽科(Anthocoridae)、姬蝽科(Nabidae),以及蝽次目(Pentatomomorpha)、黾蝽次目(Gerromorpha)、歇蝽次目(Nepomorpha)、奇蝽次目(Enicocephalomorpha)的姐妹群。奇蝽次目和黾蝽次目距离最近,与歇蝽次目共同聚合在一簇,蝽次目单独聚合为一个分支。系统发育树支持了歇蝽次目、奇蝽次目、黾蝽次目和蝽次目的单系性,而臭虫次目为并系群。

图5 基于41种昆虫线粒体蛋白质编码基因序列构建的ML系统发育树Fig. 5 Maximum likelihood phylogenetic tree of 41 insects based on the PCG sequences of mitogenome
本研究结合Illumina测序和Sanger测序,获得安康市茶网蝽线粒体基因组全长18 085 bp,是目前 NCBI登录的网蝽科昆虫线粒体全序列(15 208~16 667 bp)中长度最长的。控制区是昆虫线粒体基因组大小变异的主要来源,一般与线粒体基因组的大小呈正相关,控制区的长度差异可以发生在不同物种甚至不同个体之间[27]。目前昆虫中已报道的最短的控制区仅为70 bp,而象鼻虫的控制区长度达 9~13 kb[28]。已报道的网蝽科昆虫线粒体基因组的控制区为733~2 215 bp,而本研究中安康市茶网蝽的控制区为3 678 bp。
控制区是线粒体基因组复制和转录的主要调控区,包含线粒体基因组复制相关信息[29]。黾蝽科圆臀大黾蝽(Aquarius paludum)线粒体基因组控制区存在 TATA motif、poly T stretch、茎环结构等保守结构元件[30],地长蝽科大黑毛肩长蝽(Neolethaeus assamensis)的线粒体基因组控制区存在poly A结构、茎环结构、串联重复区等[31]。本研究发现,茶网蝽基因组中有多种重复序列和茎环结构,而重庆市茶网蝽[32]仅有 R4重复序列,没有 FER区,对于不同地域茶网蝽基因组之间的差异,有待进一步研究。
本研究茶网蝽的线粒体基因组所有区域均有明显的 AT偏好性,AT含量最高的是rRNA,这与网蝽科昆虫杨柳网蝽、菊方翅网蝽(Corythucha marmorata)、古无孔网蝽(Dictyla platyoma)中的情况相同。蛋白质编码基因的AT含量最低,这有利于物种编码其所需蛋白质的功能[33]。cox1基因是常用的DNA条形码基因,应用于昆虫的识别鉴定[34]。相对较高的 GC含量有利于降低基因的突变率,保持基因稳定性。Yang等[8]研究发现,网蝽科昆虫线粒体cox1基因的进化速率较低,atp8基因最高,本研究中安康市茶网蝽cox1基因的GC含量最高,atp8基因的GC含量最低,与其结论一致。与多数动物一样,茶网蝽线粒体基因组不同链的核苷酸组成也不对称[19]。
网蝽科昆虫线粒体基因组中绝大部分蛋白基因以ATN为起始密码子,使用最多的是ATG,其中cox1和atp6基因的起始密码子均为 ATG,ATA的使用频次略低于 ATT。终止密码子以TAA为主,cox1、atp8、nad6和nad1基因的终止密码子都是TAA;
其次为T,cox2基因均以单个T作为终止密码子,cox3和nad5基因使用 T作为终止密码子的现象较多,较少使用 TA或 TAG。昆虫线粒体基因组以 T或TA作为终止密码子现象比较常见,推测其通过转录后 mRNA多腺苷酸化形成完全终止密码子TAA[35]。
臭虫次目是异翅亚目最大的次目,有 17个科超过20 000种昆虫,其系统发育研究目前没有一致的结论。利用形态性状、转录组和核基因等研究发现,臭虫次目昆虫为单系[36-38],而使用臭虫次目昆虫线粒体基因组数据进行系统发育研究,鉴定其为并系群,尤其是网蝽科昆虫的数据,该现象更显著[8,39-40]。Liu等[39]基于线粒体基因组数据分析异翅亚目的系统发育发现,网蝽科和奇蝽次目、鞭蝽次目、黾蝽次目聚为一簇且接近发育树的根部,是其他异翅亚目昆虫的姐妹群。本研究利用线粒体PCGs进行的系统发育重建结果,能很好地支持奇蝽次目、黾蝽次目、歇蝽次目和蝽次目的单系性。但臭虫次目的情况比较复杂,其姬蝽科和花蝽科聚合在一起,形成与奇蝽次目、黾蝽次目和歇蝽次目并列的分支,而其盲蝽科和网蝽科的位置接近发育树的根部,这与Liu等[39]的研究结果相似。但本研究接近发育树根部的几个分支的支持率均低于50%,这可能与网蝽科昆虫线粒体基因组序列的变异率较大有关[8,39],较大碱基差异不容易区分系统发育关系,更适合于亲缘关系近的物种的分化检测。
猜你喜欢 控制区密码子线粒体 线粒体质量控制在缺血性脑卒中的作用研究进展医学研究生学报(2022年5期)2022-12-07靶向敲除β-珠蛋白基因座控制区增强子HS2对K562细胞转录组的影响遗传(2022年9期)2022-10-10线粒体自噬在纤维化疾病中作用的研究进展中华实用诊断与治疗杂志(2022年1期)2022-08-31镰翅羊耳蒜叶绿体基因组密码子偏好性分析福建农业学报(2021年6期)2021-08-18棘皮动物线粒体基因组研究进展海洋通报(2021年1期)2021-07-23线粒体自噬与帕金森病的研究进展生物学通报(2021年4期)2021-03-16基于OMI的船舶排放控制区SO2减排效益分析海洋通报(2020年5期)2021-01-14以“题”带“面” 突破密码子与反密码子的特性教学考试(高考生物)(2020年4期)2020-11-18密码子与反密码子的本质与拓展生物学通报(2020年11期)2020-10-22核电厂建安阶段防异物管理现代企业文化·理论版(2020年16期)2020-08-31本文来源:http://www.zhangdahai.com/shiyongfanwen/qitafanwen/2023/0714/625160.html



