【www.zhangdahai.com--其他范文】
陈 娜 邵 勤
(宜春学院生命科学与资源环境学院,江西宜春 336000)
番茄(Solanum lycopersium)为茄科茄属一年生或多年生草本植物,在我国南北各地普遍种植,具有重要的经济和营养价值[1]。番茄青枯病是由青枯菌引起的一种毁灭性的细菌性土传病害,在我国南方地区普遍发生且危害严重,每年所造成的番茄减产可达50%~100%,已成为影响番茄品质和产量的重要病害之一[2-3]。因此,加大力度研究番茄对青枯菌胁迫的响应机制,可为抗青枯病番茄品种的培育提供理论基础。
植物在其生长发育过程中为了降低各种生物与非生物逆境对其的影响,会进化出一套反应机制来抵御外界胁迫,即通过转录因子与外界信号和基因调控相整合的方式产生相应的蛋白和代谢产物来适应胁迫,从而提高植物的抗逆性[4]。NAC转录因子是近二十年来发现的植物特有的一类转录因子家族,其名称由矮牵牛(Petunia hybrida)No Apical Meristem(NAM)、拟南芥(Arabidopsis thaliana)转 录 激 活 因 子(ATAF1、ATAF2)和Cup-shaped cotyledon(CUC1/2)三个基因的首字母缩写组成[5]。通常NAC蛋白在其N-端含有高度同源且能够与DNA结合的NAC结构域,该结构域由约150个氨基酸残基组成,包含5个保守的亚结构域(A、B、C、D、E),其C-端具有转录激活功能[6]。早期研究表明,NAC转录因子广泛参与了植物的生长发育[7]、次生壁形成[8]、激素信号传导[9]、植株衰老[10]、果实成熟[11]以及各种非生物[12]和生物[13]胁迫响应过程,并且随着测序技术的迅速发展,越来越多的NAC转录因子被发掘与鉴定。如Li等[14]研究表明,葡萄(Vitis viniferaL.)VvNAC72通过抑制乙二醛酶ⅠVvGLYI-4的表达负向调控丙酮醛(methylglyoxal,MG)的解毒作用,并最终通过水杨酸介导的防御途径调节MG-相关的活性氧(reactive oxygen species,ROS)稳态,增强葡萄对霜霉病的抗性。目前有研究表明,番茄NAC转录因子也参与了植物抗病性的响应过程。Huang等[15]发现番茄SlNAC1在假单胞菌侵染过程中表达强烈上调。Liu等[16]报道番茄NAC基因SlSRN1是灰霉病菌(Botrytiscinerea)和丁香假单胞菌[Pseudomonas syringaepv.Tomato(PstDC3000)]防卫响应的正调节因子。Miao等[17]研究表明,番茄泛素连接酶SINA3(SEVEN IN ABSENTIA3)能够泛素化一个防卫相关的NAC转录因子NAC1,促进其降解,并在防卫信号转导中发挥负向调控作用。黄莹[18]在番茄中鉴定到4个能够响应番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)侵染的NAC转录因子(SlNAC20、SlNAC24、SlNAC47和SlNAC61),并通过病毒诱导的基因沉默(virus induced gene silencing,VIGS)试验得出,SlNAC61在响应TYLCV侵染时起着正向调控作用。郑晨飞[19]研究表明,番茄NAC转录因子NAC29在细菌性叶斑病抗性中具有重要的作用。Wang等[20]报道,过表达番茄NAC转录因子SlNAP1能够显著提高植株对叶斑病和青枯病的抗性。以上研究均表明,NAC转录因子在植物的抗病性调控中扮演着重要角色。但目前关于NAC转录因子调控青枯病抗性的研究较少,其调控番茄青枯病的分子机理仍不清楚。
前期,宜春学院蔬菜遗传育种与分子生物学课题组通过转录组测序及表达谱分析,筛选到一个表达差异显著的NAC转录因子SlNAP2。本研究以SlNAP2基因为研究对象,以番茄高抗青枯病和高感青枯病株系为材料,分析该转录因子的序列结构以及在不同外源激素和青枯菌胁迫下的表达模式,通过VIGS技术进一步对其参与番茄抗青枯病过程进行分析,旨在为后续深入研究NAC转录因子在番茄与青枯病互作中的作用机理提供参考依据。
1.1 试验材料
植物材料为两个番茄自交系株系,高抗青枯病材料Hm 2-2(R),高感青枯病材料BY 1-2(S),由浙南作物育种重点实验室选育得到。
番茄青枯菌病原、VIGS载体pTRV1和pTRV2菌株均由江西省作物生长和发育调控重点实验室保存,载体图谱如图1。

图1 pTRV1和pTRV2载体图谱Fig.1 The map of pTRV1 and pTRV2 vectors
pMD19-T克隆载体购自宝生物工程(大连)有限公司。大肠杆菌(Escherichia coli)感受态细胞DH5α和农杆菌感受态细胞GV3101均购自广州围谷润仪器有限公司。
1.2 番茄SlNAP2基因的克隆
利用HiPure Total RNA Mini Kit RNA提取试剂盒(Magen,广州)提取番茄高抗青枯病材料Hm 2-2的总RNA;
以HiScript II 1st Stand cDNA Synthesis Kit(+gDNA wiper)逆转录试剂盒(Vazyme,南京)进行逆转录,获得cDNA,以该cDNA为模板扩增SlNAP2基因序列。运用Primer Primer 5软件设计引物(表1),引物由苏州泓迅生物科技有限公司合成。PCR体系共20µL:10 mmol·L-1正反向引物各1µL,100~200 ng·µL-1模板cDNA 1 µL,2×Taq Master Mix(Vazyme,南京)10 µL,灭菌ddH2O 7 µL。PCR扩增程序:94℃预变性5 min,94℃变性30 s,55℃退火30 s,72℃延伸90 s,30个循环;
72℃终延伸10 min。然后将PCR产物进行电泳检测,并用Genstar胶回收试剂盒(北京康润诚业生物科技有限公司)将PCR产物进行纯化回收,最后将回收产物连接到pMD19-T载体上,用通用引物pMD19-F/R进行检测,挑取阳性克隆菌落摇菌,将菌液送至生工生物工程(上海)股份有限公司进行测序,根据测序结果确定基因的最终序列。

表1 引物序列Table 1 Primer sequences
1.3 番茄SlNAP2基因的生物信息学分析
将测序结果正确的SlNAP2基因序列通过NCBI在线软件(https://www.ncbi.nlm.nih.gov/orffinder/)确定该基因的最大开放阅读框(open reading frame,ORF),同时将该基因的核苷酸序列翻译成氨基酸序列;
运用NCBI在线程序(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)对该基因编码的蛋白结构域进行分析;
运用Clustalw2在线工具(https://www.ebi.ac.uk/Tools/msa/clustalw2/)进行同源性比对;
运用MEGA6软件进行系统进化树的构建;
运用ProtParam在线程序(https://web.expasy.org/protparam/)预测SlNAP2的蛋白理化性质;
适用Expasy-ProtScale在线程序(https://web.expasy.org/protscale/)预测SlNAP2的亲疏水性;
二级结构通过SOPMA在线程序(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测;
三级结构通过SWISS-MODEL在线程序(https://swissmodel.expasy.org/)预测;
使用TMHMM Server v.2.0在线程序(http://www.cbs.dtu.dk/services/TMHMM/)对番茄SlNAP2基因编码的蛋白跨膜结构进行预测。
1.4 番茄SlNAP2基因表达特性分析
SlNAP2组织特异性基因表达分析:在番茄长至五叶时,分别取番茄抗感青枯病材料Hm 2-2和BY 1-2的根、茎、叶片等不同组织进行液氮速冻,于-80℃条件下保存,用于后续总RNA提取。
SlNAP2在不同处理下的基因表达分析:待番茄抗感青枯病材料Hm 2-2和BY 1-2长至五叶时接种青枯菌病原(1×108CFU·mL-1),同时进行0.2 mmol·L-1水杨酸(salicylic acid,SA)和1.5 mmol·L-1茉 莉 酸 甲 酯(methyl jasmonate,MeJA)激素喷施处理,以清水喷施为对照。接种青枯菌和喷施激素处理后0、3、6、9 h取样。所有样品进行液氮速冻,于-80℃条件下保存,用于后续总RNA提取。
利用HiPure Total RNA Mini Kit RNA提取试剂盒(Magen,广州)进行总RNA提取;
以HiScript II 1st Stand cDNA Synthesis Kit(+gDNA wiper)逆转录试剂盒(Vazyme,南京)进行逆转录,获得cDNA。以番茄Actin(Solyc03g078400)作为内参基因,以StepOne Real-Time PCR仪(Applied Biosystems,Thermo Fisher,美国)进行qRT-PCR,检测SlNAP2基因在不同组织及不同处理中的相对表达量。PCR反应体系20 µL:2×RealStar Green Fast Mixture(with ROX)10µL,上下游引物qSlNAP2F/R(10 mmol·L-1)各0.4µL,cDNA 0.5µL,无RNase水补至20 µL。PCR反应程序:95℃预变性2 min;
95℃变性15 s,60℃退火/延伸20 s,40个循环;
溶解曲线由仪器自动设置。每个样品设置3个生物学重复。采用2-ΔΔCt法计算SlNAP2基因相对表达量[21]。
1.5 利用VIGS技术分析番茄SlNAP2基因功能
在已获得的SlNAP2基因cDNA序列中,选取ORF非保守区的约120 bp片段,设计引物VSlNAP2-F/R进行PCR扩增,PCR反应体系和程序同1.2。将VIGS载体pTRV2质粒和PCR扩增的回收产物分别用EcoRI和SmaI进行酶切37℃2 h,酶切后将所得产物进行酶失活70℃15 min,然后用T4连接酶[宝生物工程(大连)有限公司]将载体和PCR回收产物进行连接。连接体系10 µL:pTRV2载体2 µL,PCR回收产物5 µL,350 U·µL-1T4 DNA Ligase 0.5 µL,10×T4 DNA Ligase Buffer 2 µL,ddH2O 0.5 µL、16℃连接过夜,将连接产物采用热激法转化大肠杆菌细胞DH5α,构建TRV∷SlNAP2重组载体。将pTRV2空载体(TRV∷Empty)及重组载体(TRV∷SlNAP2)分别转化农杆菌细胞GV3101,将pTRV-1菌株分别与TRV∷SlNAP2及TRV∷Empty菌株1∶1(V/V)进行混合,注射至番茄抗病材料Hm2-2的叶片中,注射清水为对照CK。2 d后采用断根灌根法接种青枯菌,5~7 d后观察对照植株与沉默植株的表型,拍照和采样,具体步骤参照陈娜[22]的方法。提取总RNA,通过qRT-PCR测定目的基因SlNAP2的表达量,反应体系和反应程序同1.4。
2.1 番茄SlNAP2基因的cDNA序列克隆
以番茄抗青枯病Hm2-2为试验材料,提取总RNA,并反转录成cDNA,以cDNA为模板扩增SlNAP2序列,得到1 200 bp左右的扩增产物(图2-A)。切胶回收,将纯化后的片段连接至pMD19-T载体上(图2-B),挑取阳性克隆摇菌,将菌液送至生工生物工程(上海)股份有限公司测序。测序结果得出SlNAP2的全长为1 227 bp,大小与图2-A相符。
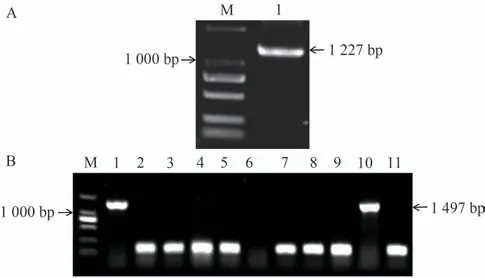
图2 番茄SlNAP2基因Fig.2 Tomato SlNAP2 gene
2.2 番茄SlNAP2基因的序列分析
进一步分析番茄SlNAP2基因序列,其ORF为828 bp(图3),起于175位,止于1 003位,共编码了275个氨基酸。在SlNAP2的N-端存在1个典型的NAC转录因子的结构域(11~135 aa)(图4)。

图3 SlNAP2 cDNA及预测的开放阅读框Fig.3 Sequences of SlNAP2 cDNA and predicted ORF

图4 SlNAP2的保守区Fig.4 Conservative domain of SlNAP2
2.3 番茄SlNAP2基因编码的蛋白同源性比对和系统进化分析
通过番茄SlNAP2编码的蛋白与其他物种的同源蛋白序列比对可知,番茄SlNAP2编码的蛋白与其他物种的蛋白相似度较高,并且SlNAP2与其他NAC转录因子一样,N-端的氨基酸序列保守性很强,可进一步划分为A、B、C、D、E 5个保守的亚结构域,C-端高度特异(图5)。运用MEGA6.0软件对番茄和其他10种植物的氨基酸序列进行聚类分析,并制作进化树,结果表明,番茄SlNAP2基因编码的蛋白与潘那利番茄SpNAP2基因编码的蛋白同源性最高,其次为马铃薯,与毛果杨、橡胶树等植物的蛋白同源性较低(图6)。

图5 番茄SlNAP2蛋白与其他物种同源蛋白多重序列比对Fig.5 Multiple alignment of homologous protein for the SlNAP2 in tomato and other species
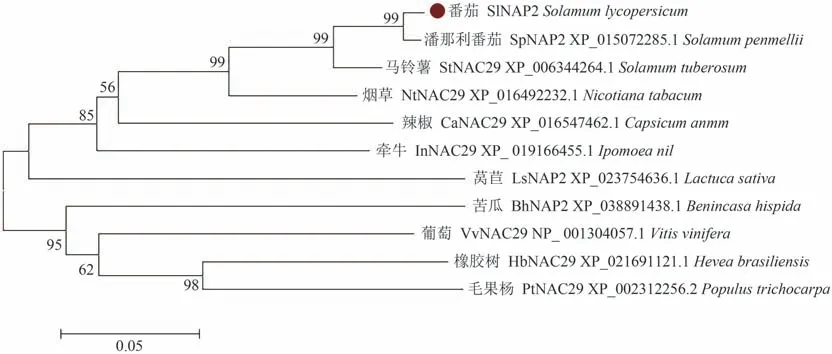
图6 SlNAP2同源蛋白的系统进化树Fig.6 Phylogenetic tree of SlNAP2 homologous protein
2.4 番茄SlNAP2基因编码的蛋白的生物信息学分析
番茄SlNAP2蛋白的理化性质用ProtParam在线程序进行分析,结果如表2所示。SlNAP2蛋白的相对分子质量为31 877.23 Da,理论等电点pI为8.56,总负电荷残基数(Asp+Glu)为31,总正电荷残基数(Arg+Lys)为34,其蛋白的分子式为C1423H2187N383O421S15,脂肪系数为62.69,不稳定指数为24.52,是一种稳定蛋白。平均亲水指数为-0.730,同时运用Expasy-ProtScale在线程序分析得出SlNAP2表现出亲水性(图7)。综上,该蛋白为碱性稳定亲水性蛋白。

表2 SlNAP2蛋白的基本理化性质Table 2 Basic physical and chemical parameters of SlNAP2 protein

图7 SlNAP2蛋白亲疏水性分析Fig.7 Hydrophilic analysis of SlNAP2 protein
2.5 番茄SlNAP2基因编码蛋白的二级结构及跨膜性预测
通过SOPMA在线程序对SlNAP2蛋白进行二级结构预测,得出α-螺旋(alpha helix)比例为21.82%,延伸链结构(extended strand)的比例为13.09%,β-折叠(beta turn)的比例为3.27%,无规则卷曲(random coil)比例为61.82%(图8-A)。以上结果表明该蛋白质的二级结构主要由α-螺旋和无规则卷曲所构成,延伸链结构和β-折叠所占的比例较低。
为进一步了解番茄SlNAP2蛋白的结构,利用SWISS-MODEL对番茄SlNAP2蛋白进行三维结构的建模分析,结果如图8-B所示。SlNAP2蛋白的主要结构是螺旋和无规则卷曲,与二级结构预测的结果一致。
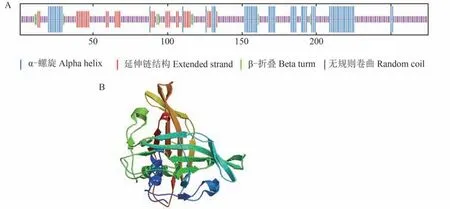
图8 SlNAP2编码的蛋白的二级结构和三级结构预测Fig.8 Secondary and tertiary structure prediction of protein encoded by SlNAP2 gene
使用TMHMM Server v.2.0在线程序对番茄SlNAP2蛋白的跨膜结构域进行分析,结果表明该蛋白不存在跨膜结构(图9)。

图9 SlNAP2基因编码蛋白的跨膜结构分析Fig.9 Transmembrane structure analysis of protein encoded by SlNAP2 gene
2.6 SlNAP2基因表达分析
2.6.1SlNAP2基因的组织特异性分析对SlNAP2在番茄不同组织中的表达进行qRT-PCR检测,结果显示,在番茄抗病材料(R)和感病材料(S)中,SlNAP2均在茎中的表达量最高,叶中次之,根中最低,并且SlNAP2在抗病材料(R)中的表达量高于在感病材料(S)中的表达量(图10-A)。表明SlNAP2在番茄中的表达具有组织特异性。
2.6.2SlNAP2基因对番茄青枯病胁迫及SA和MeJA处理的响应在番茄五叶时接种青枯菌,于不同时间点收集叶片样品进行qRT-PCR检测分析,结果如图10-B显示。接种青枯菌后,SlNAP2基因表达量不断增加,表明青枯菌能够诱导SlNAP2基因的表达,并且与番茄青枯菌侵染程度呈正相关。分别以0.2 mmol·L-1SA和1.5 mmol·L-1MeJA处理番茄植株,检测SlNAP2对外源激素的响应,结果显示,SA处理后,SlNAP2基因的表达量呈先降后升的趋势(图10-C);
MeJA处理后,SlNAP2基因的表达量呈下降的趋势(图10-D)。推测SlNAP2可能通过SA、MeJA通路参与番茄对青枯菌的胁迫响应。

图10 番茄SlNAP2基因表达分析Fig.10 Expression analysis of tomato SlNAP2 gene
2.7 SlNAP2基因对番茄青枯病的抗性调控
分别提取接种CK、TRV∷Empty和TRV∷SlNAP2后7 d的番茄叶片RNA进行qRT-PCR检测。结果显示,与接种清水和空载体TRV∷Empty的植株相比,在TRV介导SlNAP2沉默的抗病植株中,SlNAP2基因表达量显著低于CK和TRV∷Empty植株(图11-A),表明TRV∷SlNAP2接种抗病植株后能有效沉默SlNAP2基因。对番茄植株的叶片进行表型观察,接种清水和TRV∷Empty植株的叶片没有明显的变化,而接种TRV∷SlNAP2的植株叶片表现出明显的萎蔫症状(图11-B),表明沉默SlNAP2基因降低了番茄植株对青枯病的抗性。

图11 SlNAP2基因对番茄青枯病的抗性调控分析Fig.11 Analysis of SlNAP2 gene regulation of tomato resistance to bacterial wilt
NAC转录因子是植物特有的转录因子,也是植物中数量最大的转录因子家族之一。NAC转录因子不仅广泛参与植物生长发育的调控过程,而且在逆境胁迫响应过程中扮演着重要的角色,因此研究NAC转录因子的结构与功能具有重要意义。近年来,随着全球基因组测序工作的展开,很多物种的全基因组测序已经完成。到目前为止,发现甘蓝NAC基因家族包含188个基因[23],苹果为180个基因[24],水稻为151个基因[25],拟南芥为117个基因[26],毛果杨为120个基因[27],番茄为104个基因[28],葡萄为74个基因[29]。本研究鉴定了番茄NAC转录因子SlNAP2,并对其进行了生物信息学分析。序列分析结果表明,SlNAP2蛋白和其他NAC蛋白一样,其N-端含有NAM结构域,该结构域是NAC转录因子特有的,进一步可分为5个亚结构域,与大部分前人研究结果一致[30-31]。通过对SlNAP2编码的蛋白分析结果可知,该蛋白为亲水蛋白,与荣玉萍等[32]研究结果一致。蛋白质的二级结构预测可知,该蛋白以无规则卷曲为主,与谭秦亮等[33]对菠萝NAC转录因子的研究结果基本一致。
前人研究证实植物抵抗生物胁迫的防卫响应与外源激素信号分子有密切的联系,如植物激素MeJA和SA是重要的植物抗性分子,在调节植物对生物胁迫的反应中扮演着重要角色[34],而NAC转录因子作为调节植物应激响应的关键因子和植物病原菌防卫响应的重要因子而得到广泛关注。有研究证实,NAC转录因子能够通过植物激素参与植株的生物胁迫响应过程,如水稻NAC转录因子ONAC122和ONAC131在稻瘟菌(Magnaporthe grisea)侵染后诱导表达,同时ONAC122和ONAC131能够被外源激素SA和MeJA所诱导,沉默该基因提高了植株对稻瘟菌的易感性,研究结果表明ONAC122和ONAC131在水稻抗病反应中发挥重要作用[35]。Chen等[36]研究表明,茄子SmNAC通过抑制ICS1(SA合成基因)的表达来抑制SA的积累,从而降低植株对青枯病的抗性。马铃薯StNACb4能够被青枯菌、SA和MeJA所诱导,沉默NbNACb4基因可增强烟草对青枯菌的敏感性,相反过表达StNACb4基因则增强了烟草对青枯菌的耐受性,表明StNACb4在马铃薯抗青枯病胁迫中发挥关键作用[37]。本研究通过探究青枯菌和SA、MeJA激素胁迫下SlNAP2在番茄抗感材料中的表达量发现,该基因对青枯菌、茉莉酸甲酯和水杨酸胁迫处理均有不同程度的响应,表明SlNAP2确实参与响应青枯菌生物胁迫和激素胁迫。其中SA诱导的SlNAP2的表达趋势与青枯菌诱导的变化趋势相似,推测SlNAP2可能通过SA信号转导途径参与番茄对青枯菌的胁迫响应。
前人研究证实NAC转录因子参与了番茄生物胁迫的响应过程,如在烟草中过表达番茄SlNAC35能够增强植株对细菌性病原菌的抗性[38];
番茄转录因子NAC29在细菌性叶斑病抗性中具有重要作用[19];
番茄NAC转录因子NAC089是内质网应激介导的免疫(endoplasmic reticulum stress-mediated immunity,ERSI)和植物对病原体抗性所必需的[39]。本研究发现,在番茄中利用VIGS技术沉默SlNAP2之后接种青枯菌病原,番茄抗病材料的叶片表现出萎蔫症状,说明SlNAP2在番茄对青枯病的抗性中发挥着正调控作用。本研究结果证实了NAC转录因子在番茄生物胁迫响应过程中的作用,但其调控番茄青枯病的分子机理仍未清楚,今后可进一步进行SlNAP2与信号转导途径中相关基因的互作分析,并可开展SlNAP2与青枯菌病原无毒或致病基因相关蛋白互作研究,从而明确番茄SlNAP2转录因子如何调控青枯病,对SlNAP2的调控机理进行深入的分析。这对培育持久、广谱抗性的番茄品种具有重要的理论和实践意义。
本研究从番茄中克隆得到一个NAC转录因子SlNAP2,全长1 227 bp,编码275个氨基酸,为碱性稳定亲水性蛋白,且与潘那利番茄SpNAP2亲缘关系最近。qRT-PCR分析结果表明,SlNAP2基因的表达具有组织特异性,同时番茄SlNAP2基因可受到青枯菌、水杨酸和茉莉酸甲酯的诱导。进一步利用VIGS技术沉默SlNAP2后接种青枯菌,番茄植株对青枯病的抗性降低,表明SlNAP2在番茄抗青枯病过程中起正调控作用。
猜你喜欢 青枯病结构域抗性 亚洲玉米螟对Cry1Ac蛋白抗性适合度代价河北科技师范学院学报(2022年2期)2022-08-26细菌四类胞外感觉结构域的概述湖北农业科学(2022年11期)2022-07-18烟稻轮作对烟草根际土壤微生物区系及青枯病发生的影响云南农业科技(2021年6期)2021-12-30一个控制超强电离辐射抗性开关基因的研究进展空间科学学报(2021年1期)2021-05-22UBR5突变与淋巴瘤B细胞成熟实用肿瘤学杂志(2020年4期)2020-12-08甜玉米常见病害的抗性鉴定及防治中国果菜(2016年9期)2016-03-01用于黄瓜白粉病抗性鉴定的InDel标记中国蔬菜(2015年9期)2015-12-21PTPMeg2的PTP结构域对磷酸化STAT3入核的抑制作用中国老年学杂志(2015年9期)2015-01-31三种堆肥对番茄生长及青枯病防治效果的影响植物营养与肥料学报(2012年5期)2012-10-26BiologGENⅢ微孔板在烟草青枯病、黑胫病生防细菌鉴定中的应用中国烟草学报(2012年5期)2012-04-12本文来源:http://www.zhangdahai.com/shiyongfanwen/qitafanwen/2023/0724/629794.html



