【www.zhangdahai.com--其他范文】
 材料与方法
材料与方法
1.1药品与试剂
基因重组人IFN–γ:上海克隆生物高科技术有限公司,基因重组人IFN-α2b:北京远策药业有限责任公司,RPMI Medium 1640 培养基:GIBCOBRL 公司,胎牛血清:北京元亨圣马生物技术研究所, Fas/FasL单克隆鼠抗人:美国Santa cruz公司,Fas/FasL单克隆鼠抗人二抗:南京凯基生物科技发展有限公司,Annexin V-EGFP:南京凯基生物科技发展有限公司,碘化丙啶(PI):Sigma公司,MCF-7乳腺癌细胞:南京凯基生物科技发展有限公司
1.2.肿瘤细胞培养
MCF-7乳腺癌细胞以适量浓度接种于培养瓶中,加入RPMI-1640 培养液(含10%胎牛血清)置于37℃,5%CO2浓度及饱和湿度恒温细胞培养箱中培养。细胞呈单层贴壁生长,每3~4天传代一次,传代时用0.25%胰蛋白酶消化2~3min,以培养液吹打制成细胞悬液,按所需浓度接种。
1.3分组与方法
1.3.1实验分对照组和实验组,对照组即正常培养的MCF-7乳腺癌细胞,实验组为500u/ml IFN-α和500u/ml IFN-γ联合作用组。作用时间分别为24h、48h、72h。流式细胞仪检测Annexin V-EGFP染色的细胞凋亡情况、通过亚倍体峰检测PI染色细胞凋亡情况及检测处理后Fas/FasL蛋白表达情况。
1.4统计分析
所有数据资料以均数±标准差(±S)表示。采用SPSS11.5统计学软件进行统计处理,进行析因设计方差分析,两两比较用Tukey或Dunnett t检验,P<0.05表示统计学差异明显。
2.结果
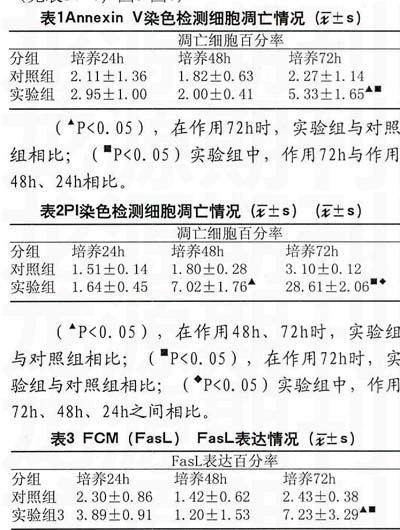
2.1流式细胞仪检测Annexin V-EGFP染色的细胞凋亡情况
随着作用时间的延长,凋亡细胞百分率有增加趋势,作用72h时,实验组与对照组比较,差异显著,有统计学意义(P<0.05);实验组中,作用72h与作用24h、48h比较,差异显著,有统计学意义(P<0.05)。(见表1,图1-图2)
2.2流式细胞仪通过亚倍体峰检测PI染色细胞凋亡情况
随着作用时间的延长,凋亡细胞百分率有增加趋势,作用48h、72h时,实验组与对照组比较,差异显著,有统计学意义(P<0.05);实验组中,作用24h、48h、72h、之间相互比较,差异显著,有统计学意义(P<0.05)。(见表2,图3-图4)
2.3流式细胞仪检测Fas/FasL蛋白表达情况
随着作用时间的延长,FasL蛋白表达有增加的趋势,作用72h时,实验组与对照组比较,差异显著,有统计学意义(P<0.05);实验组中,作用72h与作用24h、48h比较,差异显著,有统计学意义(P<0.05)。Fas蛋白表达实验组与对照组相比,差异不明显,无统计学意义(P>0.05)。(见表3、4,图5-图6)
3讨论
乳腺癌是女性最常见的恶性肿瘤,国内外乳腺癌发病率呈上升趋势,占女性肿瘤发病率的第一位,由于手术、化疗、放疗、内分泌治疗及免疫治疗等综合治疗措施的实施,乳腺癌病人的死亡率开始下降,但由于耐药、转移、复发等原因使得乳腺癌治疗的总体疗效仍然不能令人满意,因此研究探索更为有效的治疗方法显得尤为重要。
恶性肿瘤发生、发展的过程,是细胞过度增殖和细胞凋亡受到抑制的过程[1]。细胞基因调控异常,使细胞过度增殖或凋亡减弱是肿瘤的重要发生机制[2]。现代肿瘤学认为肿瘤细胞凋亡机制的失活是恶性肿瘤发生发展的普遍特性。肿瘤的发生与正常的细胞凋亡过程被抑制,破坏了细胞增殖与凋亡之间的平衡有一定关系。目前,诱导细胞凋亡被视为抗癌药物研究和开发的新靶点[3]。干扰素是一类重要的抗肿瘤、抗病毒和免疫调节作用的细胞因子。研究发现,干扰素除了增强肿瘤细胞对凋亡信号的敏感性诱导肿瘤细胞凋亡之外,还可以通过阻滞细胞周期、诱导抑癌基因的表达等作用来发挥其抗肿瘤活性。近来研究发现,干扰素还可放大由caspase诱导的线粒体功能紊乱效应,诱导细胞经线粒体途径凋亡。这些表明了干扰素的多种抗肿瘤生物学活性。
凋亡是有基因编码调控的细胞主动参与的自杀过程,以清除多余的和衰老的或损伤的细胞。凋亡主要被一个天冬氨酸特异的半胱氨酸蛋白酶(aspartate specific cysteineproteases)家族,即半胱天冬蛋白酶caspases来执行。目前公认的三条凋亡途径:(1)死亡受体途径。(2)线粒体途径。(3)内质网途径。研究发现在哺乳动物细胞中主要存在两种细胞凋亡途径即死亡受体途径[4]和线粒体途径[5,6],他们通过一系列分子和生物化学途径导致两条途径的共同的“中央处理器”分子即caspases的活化。其中死亡受体途径:死亡受体与特异性配体结合而发生自聚化,即而通过转接器蛋白募集、激活肤氨酸门冬氨酸蛋白酶(Caspase)-8,继而激活下游procaspase,引起caspase级联效应,破坏DNA完整性,干扰细胞周期正常运行,破坏细胞结构而最终导致典型的细胞凋亡形态改变[7]。线粒体途径:即内源性凋亡途径,是细胞外界的信号以及DNA损伤通过Bcl-2家族提高线粒体膜的通透性,使线粒体释放出促进凋亡的细胞色素C(cytC),cytC通过与接头蛋白Apaf-1(apoptosis activating factor-1)发生寡聚化而与半胱天冬氨酸蛋白酶原(procaspase)procaspase-9形成凋亡小体,激活caspase-9,进而激活“效应”caspases(如caspase-3),随之活化多种核酸内切酶(Dnase)裂解细胞内的关键蛋白,从而使细胞产生凋亡形态学改变,导致细胞凋亡[8]。通过调控细胞信号转导系统诱导细胞凋亡以治疗肿瘤正成为目前肿瘤学领域的研究热点之一,在调控细胞凋亡的众多信号转导系统中,Fas/FasL介导的通路是引起细胞凋亡的重要途径之一,Fas和FasL均主要以膜形式存在,两者结合可引起多种细胞凋亡,大多数肿瘤细胞都同时表达Fas和FasL, 在机体抗肿瘤的免疫反应中Fas和FasL途径尤为重要。然而由于肿瘤患者这一途径发生了变化,存在比较普遍的Fas和FasL系统的异常,如Fas表达下降和FasL异常表达。IFN-γ可以通过干扰素受体上调肿瘤细胞的Fas蛋白和淋巴细胞的FasL从而加强这一肿瘤免疫途径[9]。临床应用的许多治疗方法的作用机制之一是通过影响Fas和FasL引发肿瘤细胞凋亡。Fas配体(Fas ligand,FasL)和死亡受体(Fas)结合是介导细胞凋亡的最主要途径之一,Fas又称Apo-1或CD95,属于TNFR(tumor necrosis factor receptor)家族,FasL分子胞外区与TNF高度同源,FasL蛋白为Fas的体内天然配体[10,11],FasL与Fas结合介导细胞凋亡,故FasL又称死亡因子,FasL与Fas结合后,Fas相关死亡区蛋白(FADD)激活Caspase,组成死亡诱导信号复合(DISC),pro-caspase-8(-10)可自身催化成活性异四聚体形式[12],继而激活下游procaspase,再激活一系列caspase1、3、7等级联反应促使Fas蛋白所在的细胞的凋亡发生[13-15]。
研究发现,Ⅰ型和Ⅱ型干扰素都具有调节和增强多数肿瘤细胞对Fas介导的凋亡敏感性的作用[16]。IFN-α可通过直接活化Caspase-3而激活caspase-8,或通过使线粒体释放细胞色素C而间接活化caspase-3诱导细胞凋亡信号的转导,调节肿瘤细胞对Fas介导细胞凋亡的敏感性[17]。Liedtke等[18],报道IFN-α能上调肝癌细胞的caspase-8和Fas的表达来介导其凋亡。IFN-γ与其相应受体结合后首先活化非受体型酪氨酸激酶,继而导致转录激活因子1(signal transductors and activator of transcription 1,STAT1)磷酸化和二聚化,STAT1与位于干扰素调节因子1(interferon regulatory factor-1,IRF-1)启动子区域的IFN-γ活化位点紧密结合,激活IRF-1基因的表达。由于caspase-3和caspase-7基因是IRF-1作用的重要靶基因,特别是caspase-7是诱导肿瘤细胞凋亡的关键分子,在抑制肿瘤细胞bcl-2的抗凋亡效应和诱导凋亡过程中发挥关键作用[19]。Inaba等[20]报道INF-γ有增强由Fas/FasL介导的骨肉瘤细胞凋亡作用。本实验结果可见,两种干扰素联合应用可诱导MCF-7乳腺癌细胞凋亡,随着作用时间的延长,凋亡细胞百分率有增加趋势;也可提高细胞FasL蛋白的表达,随着处理时间的延长,FasL蛋白表达百分率有增加的趋势,差异有统计学意义(P<0.05),干扰素对MCF-7乳腺癌细胞Fas蛋白的表达无明显影响。可见两种干扰素联合应用可能通过增加MCF-7乳腺癌细胞FasL蛋白的表达,进而增加Fas/FasL的交联,增加Fas/FasL通路的敏感性来诱导MCF-7乳腺癌细胞的凋亡。研究干扰素诱导肿瘤细胞凋亡的多种机制和途径,将对干扰素的临床应用有很大的指导意义。
结论
干扰素-γ、干扰素-α2b二者联合应用可诱导MCF-7乳腺癌细胞凋亡,作用机制可能与通过上调FasL蛋白的表达,加强Fas/FasL之间的交联及使Fas/FasL通路敏感性增加有关。
[1] Meyn RE,Stephens LC,Hunter NR,et al.Kinetics of cisplatin-induced apoptosis in murine mammary and ovarian adenocarcinomas[J].Int J Cancer,1995,60(5):725-729.
[2] Ssholz M,Cinatl J.Fas/FasL interaction :anovelimmune the rapy approach with immobilized biologicals [J].Med Res Rev,2005,25(2): 331-342.
[3] Nicholson DW.From bench to clinic with apoptosis based therapeutic agents[J].Nature,2000,407(6805):810-816.
[4] Srivastava RK.Intracellular mechanisms of TRAIL and its role in cancer therapy[J].Mol Cell Biol Res Commun,2000,4(2):67-75.
[5] Kroemer G. Cytochrome c[J].Curr Biol,1999,9(13):468-481.
[6] Du C,Fang M,Li Y,et al.Smac,a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition[J].Cell,2000,102(1):33-42.
[7] Arita K,Utsumi T,Kato A,et al.Mechanism of dibucaine-induced apoptosis in promyelocytic leukemia cells(HL-60)[J].Biochem Pharmacol, 2000,60(7):905-915.
[8] Samuni AM,Kasid U,Chuang EY,et al.Effects of hypoxia on radiation-responsive stress-activated protein kinase,p53,and caspase 3 signals in TK6 human lymphoblastoid cells[J].Cancer Res,2005,65(2): 579-586.
[9] Nagao M,Nakajima Y,Kanehiro H,et al.The impact of interferon gamma receptor expression on the mechanism of escape from host immune surveillance in hepatocelluar carcinoma[J].Hepatol,2000,32(3):491-500.
[10] Zhu Q,Liu JY,Xu HW,et al.Mechanism of counterattack of colorectal cancer cell by Fas/Fas ligand system[J].World J Gastroenterol,2005, 11(39):6125-6129.
[11] Abrams SL.Positive and negative consequences of Fas/Fas ligand interactions in the antitumor response [J].Front Biosci,2005,10(4): 800-821.
[12] Shi Y.Capsize activation:revisiting the induced proximity model [J].Cell,2004,117(7):855-858.
[13] Kamei T,Inui M,Nakase M,et al.Experimental therapy using interferon-gamma and anti-Fas antibody against oral m alignant melanoma cells[J].Melanoma Res,2005,15(5):393-400.
[14] Nomura J,Matsumoto K,Iguchi-Ariga SM, et al. Mitochondria-independent induction of Fas-mediated apoptosis by MSSP[J].Oncol Rep,2005,14(7):1305-1309.
[15] Grassme H,Kirschnek S,Riethmueller J,et al.CD95/CD95 ligand interactions on epithelial cells in host defense to pseudomonas aeruginosa[J].Science,2000,290(5491):527-530.
[16] Kurdi M,Booz GW.Jak inhibition,but not Statl knockdown,blocks the synergistic effect of IFN-γ on Fas-induced apoptosis of A549 human non-small cell lung cancer cells[J].J Interferon Cytokine Res,2007,27(1): 23-31.
[17] Kelly JD,Dai J,Eschwege P,et al.Dowegulation of Bcl-2 sensitises interferon-resistant renal cancer cells to Fas[J].Br J Cancer,2004, 91(1):164-170.
[18] Liedtke C,Groger N,Manns MP,et al.Interferon-alpha enhances TRAIL-mediated apoptosis by up-regulating caspase-8 transcription in human hepatoma cells[J].J Hepatol,2006,44(2):342-349.
[19] Song RX,Zhang Z, Mor G,et al.Down-regulation of Bcl-2 enhances estrogen apoptotic action in long-term estradiol-depleted ER(+) breast cancer cells[J].Apoptosis,2005,10(3):667-678.
[20] Inaba H,Glibetic M,Buck S,et al.Interferon-gamma sensitizes osteosarcoma cells to Fas-induced apoptosis by up-regulating Fas receptors and caspase-8[J].Pediatr Blood Cancer,2004,43(7):729-736.
本文来源:http://www.zhangdahai.com/shiyongfanwen/qitafanwen/2023/0401/578170.html



