【www.zhangdahai.com--其他范文】
马瑞乡 黄满昌 张佳佳 赵澳舜 丁兴萃 罗自生 刘胜辉 肖子璋 沈 凯
(1.国家林业和草原局竹子研究开发中心 浙江省竹子高效加工重点实验室 杭州 310012;
2.南京林业大学 南京 210037;
3.浙江大学生物系统工程与食品科学学院 杭州 310058)
毛竹(Phyllostachysedulis)为笋材两用散生竹种,是世界上最重要的经济竹种之一。毛竹笋生长发育具有两大特点:生长快,老化快(郑笑等,2009;
王杰,2016)。竹笋快速生长是每个竹节间分布居间分生组织的细胞分裂、增大、伸长(Longetal., 2017; 熊文愈等, 1980; 于芬, 2008)、基因调控(Pengetal., 2013a; 2013b; Yehetal., 2013)、激素调控(丁兴萃, 1997)、酶调控(Cuietal., 2012)等综合调控作用的结果,竹笋老化是激素调控(罗自生, 2006)、活性氧代谢(席屿芳, 2001)、酶调控(罗自生, 2006; 王敬文, 2002)等综合调控作用的结果。
植物呼吸作用分为无氧呼吸和有氧呼吸2种,无氧呼吸主要发生在种子萌发初期、块茎、果实内部生长发育阶段(武维华, 2018);
有氧呼吸主要包括糖酵解(glycolysis, EMP)、三羧酸循环(tricarboxylic acid cycle, TCA)和戊糖磷酸(pentose phosphate pathway, PPP)等代谢途径(Fernieetal., 2004; Heidarvandetal., 2017; 赵明新等, 2012),每条途径均经过一系列酶促反应,其中丙酮酸激酶(pyruvate kinase, PK)、琥珀酸脱氢酶(succinate dehydrogenase, SDH)、葡萄糖-6-磷酸脱氢酶(glucose 6-phosphatedehydrogenase, G-6-PDH)与6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehydrogenase, 6-PGDH)分别是糖酵解(EMP)、三羧酸循环(TCA cycle)和戊糖磷酸途径(PPP)的关键限速酶(Linetal., 2019; 2020; 黄鹏云等, 2009; Niehausetal., 2020),烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide, NAD)、还原型烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide reduced, NADH)以及烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate, NADP)和还原型烟酰胺腺嘌呤二核苷酸磷酸(Nicotinamide adenine dinucleotide phosphate, NADPH)含量变化分别反映EMP、TCA以及PPP代谢情况(Galeazzietal., 2011; Grahametal., 2009; 黄骥等, 2004)。PPP与植物受多种环境胁迫应答紧密相关,在种子萌发(陈丽培等, 2012)水涝(彭克勤等, 2001)、低温(林善枝等, 2001)和病原体侵染(Indeletal., 2002)等胁迫下,G-6-PDH和6-PGDH的活性或基因表达水平均显著增强。线粒体是细胞内的“动力工厂”,为细胞活动提供能量(翟中和, 2000),线粒体内嵴数量(卫星等, 2010)、线粒体数目(陈丽英等, 2015)和分布(Zhengetal., 2009)随其生理状态和外界环境而变化,线粒体的形态在植物凋亡早期可维持正常功能,但随着凋亡过程会相继出现肿胀、空泡化等一些特异性的变化(李娟, 2009)。
本研究以未出土毛竹笋为对象,探究毛竹笋采后在空气和高浓度氧气环境下储存过程中笋体的线粒体、氧气浓度、呼吸强度、呼吸代谢途径关键酶活性和代谢底物与产物等的变化,旨在揭示毛竹笋采后呼吸作用和老化机制,为竹笋保鲜提供理论依据。
研究区位于浙江省湖州市吴兴区( 30°48′N, 119°59′E),海拔30 m,属北亚热带气候区,四季分明,雨热同季,年均气温15.4 ℃,最高气温30.8 ℃,最低气温-0.5 ℃,年均降水量1 243 mm。
2.1 试验材料
2021年3月22日,选取未出土(长度15~20 cm,直径6~8 cm)和出土3 m高竹笋,竹笋无机械损伤、无虫害且大小一致。毛竹笋挖掘后,置于泡沫箱内,上层放置干冰并用多层硬纸板隔开,2 h内运送至实验室进行处理。
2.2 试验方法
2.2.1 分组处理 竹笋先用清水清洗干净,再用1%次氯酸钠溶液浸泡杀菌30 s后取出晾干,随机分为2组:
一组置于常温密闭玻璃容器内用高浓度氧气(90%O2+10%N2)处理,每天每隔8 h注入气体1次,每次30 min,注气后密封保存,另一组置于常温空气(21%O2+79%N)中作为对照,2组处理同时放置盛有足够水和1 mol·L-1的NaOH溶液的烧杯,保证湿度和吸收二氧化碳,并用温、湿度计监测温、湿度变化,保证温度为(20±1) ℃,空气湿度在95%以上。每隔2天取1次样,每次取样随机取6株竹笋作为6个生物学重复,每株竹笋分上下部分取样(从竹鞭与竹笋连接处的“螺丝钉”往笋尖数第5节位置作为下部取样,第10节位置作为上部取样),每次取样时采用氧电极检测玻璃容器内氧气浓度,样品保存至-80 ℃冰箱,用于呼吸代谢过程相关指标测定。在同一样地随机选取6株(3.0±0.1) m高出土毛竹笋,取出土部位向笋尖方向第0、8、16、24节,未出土毛竹笋从“螺丝钉”向笋尖第5、10节各取1 cm3笋块,用于切片观察。
2.2.2 腐烂指数统计 腐烂指数参照以往研究方法(Caoetal., 2013; Jiaoetal., 2018; Shenetal., 2021; 陈存坤等, 2016)计算。腐烂等级分为5级:0级切面未发生腐烂或颜色加深;
1级切面略微开始腐烂变色但面积不超过切面的1/2;
2级切面腐烂面积超过1/2且腐烂变色加深;
3级切面完全腐烂变色且颜色为深褐色;
4级切面完全腐烂变为深褐色且笋体变软。每次取样随机取6株竹笋,记录从竹笋“螺丝钉”至笋尖第10节切面腐烂情况,腐烂指数计算公式如下:
式中:X为腐烂指数;
Rn为腐烂等级;
n为该等级竹笋个数;
nmax为总竹笋数;
kmax为最高等级数。
2.2.3 呼吸速率测定 在两试验组内,每次分别随机选取10株毛竹笋,使用GXH-3010E型红外二氧化碳分析仪测定呼吸速率(Chenetal., 2020; Linetal., 2019; 2020; 陈梦茵等, 2015),结果以CO2变化率计算。
2.2.4 玻璃容器和笋体内氧浓度测定 采用氧电极(Unisense OX-NP, 丹麦)探针,分别插入密闭玻璃容器内及竹笋第5、10节横切面中心和边缘1 cm深度,稳定20 s后测定。每个测定点3次重复,计算玻璃容器和笋体氧含量(Greveetal., 2003; Wilbanksetal., 2014; Xiaoetal., 2018)。
2.2.5 有氧呼吸占比计算 测定密闭玻璃容器内12 h的O2消耗量,计算对应CO2量即为有氧呼吸CO2生成量,用有氧呼吸CO2生成量与红外二氧化碳分析仪测得CO2生成量的比值表示有氧呼吸占比,计算公式如下:
式中:a为有氧呼吸占比;
CO2为O浓度变化量(mg·L-1);CCO2为CO2浓度变化量(mg·L-1);
V为容器体积。
2.2.6 线粒体结构观察 将1 cm3笋块用FAA(50%无水乙醇∶冰醋酸∶甲醛=18∶1∶ 1)固定,经双固定、脱水、渗透、包埋、染色、切片和定位等步骤,在Hitachi H-7650型透射电镜下观察竹笋细胞组织和线粒体结构。
2.2.7 酶活测定 取试样竹笋上下部位各10 g,分别测定呼吸代谢过程相关酶活性。SDH酶活性测定参照Li 等(2016)和 史青纯(2009)方法;
PK酶活性测定参照Burrell等(1994)和高尚(2021)方法;
G-6-PDH和6-PGDH活性测定参照Lin等(2020)方法。
2.2.8 NAD、NADH、NADP、NADPH和乙醇含量测定 采用高效液相色谱(HPLC)测定NAD、 NADH、NADP、NADPH和乙醇的量。取10 g制备好的样品匀浆于100 mL具塞离心管中,加入20.0 mL乙腈,4 000 r·min-1离心2 min,加入约5 g氯化钠,剧烈振摇1 min,4 000 r·min-1离心5 min,使乙腈和水分层。取10 mL乙腈层于50 mL梨形瓶中,40 ℃下旋转蒸发浓缩至近干,洗耳球吹干后用溶解残渣待净化,供超高效液相色谱测定。色谱参数设置具体如下:
流动相流速 0.5 mL·min-1;
荧光检测波长Ex=280 nm,Em=330 nm;
柱温 40 ℃;
进样量 4 μL。取NAD、NADH、NADP、NADPH和乙醇标准品(纯度>99%)用于定量测定。
2.3 统计分析
获得的氧浓度、呼吸速率、酶活性和物质含量数据进行显著性和差异性方差分析,选用三倍方差剔除法剔除异常值,分析结果用Origin 2022软件绘图。
3.1 竹笋采后储存过程中腐烂情况
采后2天,竹笋仍处于新鲜状态,无腐烂情况发生。随着放置时间延长,竹笋开始出现腐烂,4~6天腐烂最快,6天时笋体已明显腐烂。空气处理组的竹笋腐烂程度和速度高于高氧处理组(图1),空气处理组第2天开始出现腐烂,第4天的腐烂指数是高氧处理组的2倍,4~6天两处理组的腐烂指数均剧增(表1),这表明高氧环境能够延缓采后竹笋开始腐烂的时间和程度。

表1 采后毛竹笋腐烂指数Tab.1 Perishable index of postharvested moso bamboo shoot
3.2 竹笋超微结构观察
由图2可知,竹笋快速生长阶段,笋体上部与下部线粒体数量差异不大,线粒体形态完整且以圆形为主,第0节(图2a)和第8节(图2b)线粒体颜色更深,结构更致密,表明其内嵴数量较多;
第16节(图2c2)细胞线粒体中部观察到空泡和线粒体肿胀现象,且线粒体内嵴数量较少,这与细胞凋亡相关,可能是由于毛竹笋快速生长阶段能量供应需要通过细胞自噬实现;
第24节为笋尖位置(图2d、d1、d2),细胞内基质丰富,细胞活动旺盛;
笋下部细胞明显较上部细胞大,细胞壁加厚明显,表明竹笋快速生长阶段期上部细胞以分裂为主,下部细胞以增大、伸长和细胞壁增厚为主。线粒体在竹笋整个笋体大量分布,通过有氧呼吸产生能量供应竹笋旺盛生命活动。
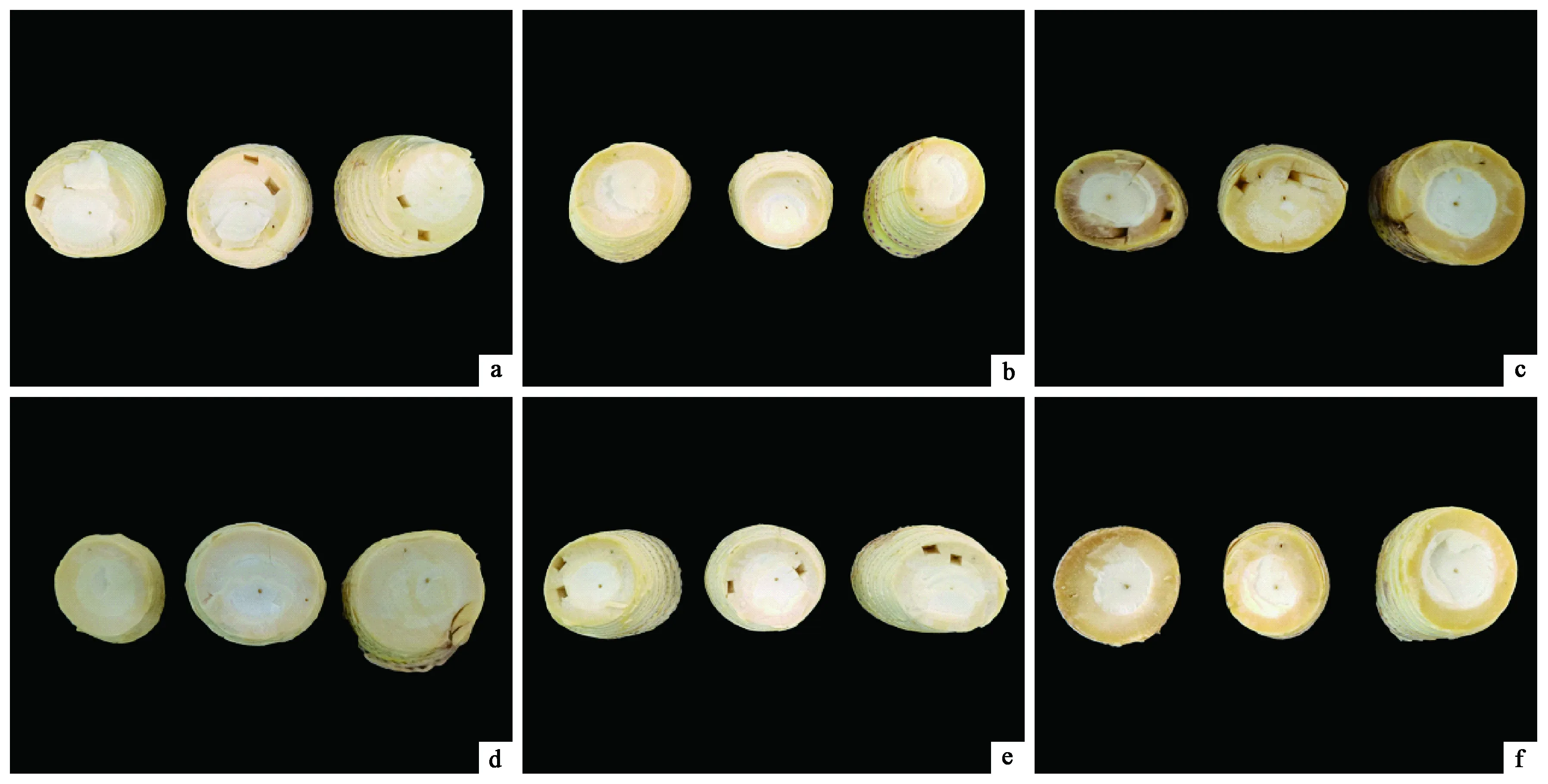
图1 竹笋采收后储存过程中腐烂情况观察Fig. 1 Observation on the decay of postharvested moso bamboo shoot during storagea、b、c为空气处理2、4、6天切面;
d、e、f为高氧处理2、4、6天切面。a, b and c are the sections treated by atmosphere for 2, 4 and 6 d; d, e and f are sections treated by high-concentration oxygen for 2, 4 and 6 d.

图2 竹笋快速生长阶段超微结构观察Fig. 2 Ultrastructure observation of moso bamboo shoot during rapid growth period红色箭头指向为线粒体,a、b、c、d为毛竹笋出土3 m高时第0节、第8节、第16节、第24节的组织结构,a、a1、a2表示毛竹笋出土3 m高时第0节不同倍数的超微组织结构(3 000×、10 000×、30 000×),b、c、d以此类推。The red arrow points to mitochondria; a,b, c and d are the tissue structure of segment 0, 8, 16, and 24 when moso bamboo shoots were excavated at 3 m high; a, a1, and a2 are the ultrastructure (3 000×, 10 000×, 30 000×) of segment 0 at different times when bamboo shoots were excavated at 3 m high; b, c and d are the same.

图4 竹笋采后储存过程中笋体内氧气含量和呼吸速率变化Fig. 4 Changes of oxygen content and respiration rate in post-tharvested moso bamboo shoot during storage不同小写字母表示不同时间点空气处理存在显著差异(P<0.05),不同大写字母表示不同时间点高氧处理存在显著差异(P<0.05),*表示同一时间两处理之间存在显著差异,误差线表示标准误。下同。Different lowercase letters represent significant difference between aotmosphere treatments at different time points (P < 0.05), different uppercase letters represent significant difference between high-concentration oxygen treatments at different time points (P < 0.05), * represents significant difference between two treatments at the same time, and the error bar represents standard error, the same below.
由图3可知,空气处理0天时竹笋薄壁细胞内含物丰富,细胞内线粒体完好且数量较多(图3a、a1、b、b1),2天时细胞内含物明显减少,4天时观察到线粒体结构有空泡;
高氧处理线粒体较空气处理小,但数量分布较多,6天时高氧处理组线粒体数量明显较空气处理多(图3k、l、m、n),表明高氧处理可延缓竹笋线粒体解体,高氧环境可能通过延缓线粒体解体从而提高竹笋有氧呼吸能力。空气和高氧2种处理的竹笋组织结构在空间上具有相似性:
上部可以明显观察到细胞核且数量多于下部,下部细胞体积较上部大,且细胞壁出现增厚现象,这与竹笋的生长特性有关,竹笋是自下而上逐渐生长成熟的,上部分生组织细胞分裂分化,而下部组织日趋成熟(Wangetal., 2019; 孙小青, 2014)。笋体内线粒体形状以圆形和椭圆形为主。从未出土阶段到快速生长阶段,竹笋线粒体分布从上部多、下部少转变为整个笋体数量均增多,且下部线粒体内嵴数量增多。出土竹笋细胞壁在储存后期会出现增厚现象,而快速生长阶段竹笋下部细胞的细胞壁有明显增厚现象,这是竹笋木质化增强等因素导致其老化造成的。
3.3 竹笋呼吸速率与氧含量的变化
在空气和高氧2种环境下,竹笋采后前4天笋体氧含量无显著差异且均处于较低水平,第4~6天笋体氧含量显著增加,其变化趋势与其腐烂速度相吻合,6天时高氧处理组笋体氧含量至3.57 mg·L-1,空气处理组笋体氧含量增至2.08 mg·L-1,高氧处理组笋体氧含量约为空气处理组的1.5倍(图4)。
空气处理竹笋呼吸速率随着时间延长呈逐渐降低趋势。高氧处理竹笋呼吸速率显著高于空气处理,呈先增后降趋势:
2天时达到呼吸峰值(1 287.89 mgCO2·kg-1h-1),是空气处理呼吸速率的4.45倍;
6天时处理,空气组呼吸速率为24.01 mgCO2·kg-1h-1,高氧处理组呼吸速率为168.55 mgCO2·kg-1h-1,是空气处理组的7.02倍;
空气组呼吸速率6天时约为0天时的1/20,高氧处理组6天时约为0天时的1/3(图4)。呼吸速率变化与线粒体分布吻合,表明高氧浓度环境可促进采后竹笋的有氧呼吸强度。
第2天时空气处理有氧呼吸占比为40.39%,高氧处理有氧呼吸占比为95.92%,表明竹笋储存过程中有氧呼吸与无氧呼吸共存,空气处理下以无氧呼吸为主,高氧处理下以有氧呼吸为主。
3.4 竹笋酶活性变化
空气处理竹笋PK、SDH、6-PGDH和G-6-PDH等酶活性均呈增加趋势,其中SDH活性增加最显著,第6天时竹笋上部SDH活性增至348.41 nmol·min-1mg-1prot,是0天时的4.77倍,竹笋下部SDH活性增毛262.33 nmol·min-1mg-1prot,是0天时的6.73倍(图5);
6-PGDH和G-6-PDH活性之和呈增加趋势,且上部酶活性高于下部。与空气处理相比,高氧处理竹笋PK、SDH、6-GPDH、G-6-PDH活性显著增加,SDH、6-GPDH和G-6-PDH活性均为前4天增加且达到峰值之后减少。呼吸作用关键酶活性变化趋势表明,竹笋在空间上存在上部较下部呼吸旺盛的现象,各呼吸途径关键酶活性均呈增加趋势,其中随着储存时间延长,TCA循环的关键酶活性增强趋势最为显著。高氧处理EMP、TCA循环和PPP的关键酶活性增强显著提高了呼吸速率。

图5 竹笋储存过程中笋体呼吸作用相关酶活性Fig. 5 Changes of respiration related enzyme activities in moso bamboo shoot during storagea、c、e表示竹笋上部,b、d、f表示竹笋下部。a, c and e represent the upper part of bamboo shoot, and b, d and f represent the lower part of bamboo shoot.
3.5 竹笋NAD、NADH、NADP、NADPH和乙醇含量变化
NAD和NADP是EMP和TCA循环的底物和产物,NADP和NADPH是PPP的底物和产物。由图6可知,空气处理0~2天时NAD含量增加,NADH含量变化并不显著,NADP含量减少,NADPH含量增加,表明竹笋呼吸作用主要以PPP途径为主;
2~6天过程中NAD含量显著减少,NADH含量显著增加(P<0.05),NADP含量显著减少,NADPH含量变化不显著,表明竹笋呼吸作用以EMP含量和TCA为主。与空气处理相比,高氧处理第4天时NAD含量增加,NADP含量没有显著变化,2~4天时NADPH含量显著降低,4~6天时NADH含量显著降低,表明高氧处理促进了PPP代谢。
乙醇是无氧呼吸的产物,空气和高氧处理竹笋体内乙醇含量均呈慢—快—慢的增加趋势,二者含量均无显著差异。空气处理6天时竹笋上部乙醇含量比0天的增加48%,下部增加42.7%,其中2~4天乙醇含量增加最快。高氧处理未显著降低竹笋体内乙醇含量,只有4天时竹笋上部乙醇含量显著比空气处理低(P<0.05)。以上结果表明,储存过程中竹笋无氧呼吸产生乙醇一直处于累加状态,高氧储存环境并不能有效遏制减弱无氧呼吸。
4.1 竹笋呼吸途径变化
植物在干旱、淹水或盐胁迫等情况下,呼吸作用呈现PPP增强(刘晓忠等, 1992; 彭克勤等, 2001; 张晓婷等, 2008; 赵欣等, 2018)。本研究中,采后竹笋置于空气中时,呼吸作用以无氧呼吸为主、有氧呼吸为辅,且EMP、TCA和PPP 3种途径交替进行,其中PPP最强阶段为0~2天,说明采后竹笋处于胁迫状态,导致PPP暂时增强。第4~6天,竹笋离体时间过久,虽然EMP、TCA循环和PPP 3种呼吸途径关键酶活性未降低,但底物NAD和NADP含量已降至最低,产物NADH、NADPH和乙醇含量均无显著变化,线粒体出现肿胀和空泡现象(图3g、k、m),研究发现拟南芥(Arabidopsisthaliana)线粒体空泡和肿胀与细胞死亡有密切关系(李娟, 2009),因此总体呼吸速率显著降低(图4)。空气处理组竹笋在整个储存期间呼吸速率呈降低趋势,初始时最高,可能是采后的创伤应激激活了一系列防御反应(谢露露等, 2020),增加了活性氧含量(Schilmilleretal.,2005),短时间刺激使呼吸速率增加,之后逐渐降低。

图6 竹笋采后储存期间笋体NAD、NADH、NADP、NADPH和乙醇含量变化Fig. 6 Changes of NAD, NADH, NADP, NADPH and ethanol contents in post-harvested moso bamboo shoot during storagea、c、e、g、i表示竹笋上部,b、d、f、h、j表示竹笋下部。a,c, e, g and i represent the upper part of bamboo shoots, and b, d, f, h and j represent the lower part of bamboo shoots.
4.2 竹笋生长发育过程中呼吸作用的供能物质
呼吸作用可为生物体内蛋白质、脂肪和多糖等物质合成提供能量和原料,与生长发育关系密切(于超, 2021)。Cui等(2012)研究表明,在笋箨紧包笋体阶段,竹笋的生长发育、光合作用能力很弱,属于异养型,其营养物质主要通过贮藏或新合成的脂类物质通过β-氧化和乙醛酸循环转化为糖,满足在糖分不足情况下竹笋生长发育的能量和物质需要。从冬季休眠到出土后1.5 m快速生长的竹鞭—竹笋生长系统中,淀粉主要储存在竹笋笋体,淀粉水解形成可溶性糖直接供应生长发育,竹鞭为淀粉运输的“通道”,而非原来认为通过储存在竹鞭的淀粉水解成可溶性糖运输至笋体供应生长发育(Zhangetal., 2021; 张佳佳等, 2021)。本研究超微组织中观察到笋体内分布大量淀粉粒且空间分布呈现上部较下部多的现象,与线粒体的空间分布一致。竹笋采后第0天切片中观察到淀粉粒分布,但到第6天呼吸停止时超微切片中无法观察到淀粉粒,表明随着储存时间延长,淀粉被呼吸作用消耗产生能量和其他代谢物质,同时EMP、TCA循环和PPP的底物NAD和NADP含量也为最低水平。但竹笋脂质含量很低(江泽慧, 2002),难以满足笋体较强呼吸作用产生能量的需要,因此,竹笋采后呼吸作用的主要供能物质是糖类,而非脂类(Cuietal., 2012)。
4.3 高氧处理竹笋TCA循环和PPP途径与其腐烂的关系
高氧处理可显著提高采后竹笋的呼吸速率,增强TCA循环和PPP,表明高氧处理能够显著增加竹笋有氧呼吸能力,延缓采后竹笋腐烂。高氧处理可显著降低NADH和NADPH含量,NADPH含量降低反馈促进了6-GPDH和G-6-PDH活性增加,从而促进PPP途径,NADH含量降低也反馈促进了TCA循环的进行(Maynardetal., 2020),同时G-6-PDH和6-PGDH活性之和与SDH活性均显著增强,乙醇含量降低,表明高氧处理主要通过TCA循环和PPP增强促进呼吸作用,抑制无氧呼吸。高氧处理笋体下部呼吸相关酶活性呈显著增加趋势,表明高氧处理可显著增加竹笋下部呼吸途径相关酶活性,即高氧处理使竹笋从原来上部为活动中心转变为整株笋体均处在活跃的代谢状态,是竹笋储存过程中应对外界氧含量剧增的应激反应,旨在维持自身高的呼吸速率,持续为竹笋生命活动提供能量。
目前,竹笋保鲜主要采用低温结合其他技术方法,如带壳毛竹笋25 ℃放置9天中完全腐烂,在4 ℃下用臭氧结合气调方法储存,28天后腐烂率为15%(张慧, 2018),本研究中高氧处理有效延缓了采后竹笋的腐烂速率,在草莓(Fragaria×ananassa)、白菜(Brassicacampestrisssp.chinensis)等果蔬中也得到相同结果(Luetal., 2018; Wangetal., 2020; Zhangetal., 2015)。苹果(Maluspumila)和梨(Pyruspyrifolia)采后,当能量供应不足时褐变腐烂加剧(Saquetetal., 2000),而当处于高能量状态时细胞可以通过活性氧清除酶或非酶系统抗氧化剂作用清除活性氧维持细胞膜功能,延缓衰老和腐烂(Yietal., 2008;
2010)。高氧环境下,采后竹笋呼吸速率提高,诱发氧化应激延缓竹笋腐烂,该结果与高氧环境刺激植物组织进行高呼吸速率吻合(Jacxsensetal., 2001)。Ghidelli等(2015) 对洋蓟(Cynarascolymus)的研究也有同样结果。龙眼(Dimocarpuslongan)、番茄(Lycopersiconesculentum)和西兰花(Brassicaoleraceavar.botrytis)在储存过程中PPP增强对延缓竹笋腐烂有积极作用(Guoetal., 2014; Linetal., 2019; Linetal., 2017; 陈梦茵等, 2015),高氧处理增加竹笋PPP,同样在空气处理中,PPP所占比例降低腐烂加剧,即采后前2天的呼吸途径以PPP为主时,竹笋腐烂指数仅为4.17%;
2~4天,PPP比例降低,笋体腐烂指数增至16.67%;
4~6天,PPP几乎停止,竹笋腐烂指数达62.5%,表明竹笋腐烂受PPP代谢途径强弱的调节,PPP代谢减弱会加速竹笋腐烂,也证实高氧处理通过增加PPP代谢强度而延缓毛竹笋腐烂。无氧呼吸产物乙醇积累后会进行乙醇发酵,导致采后蔬果腐烂(Alietal., 2021; Jamesetal., 2000),高氧处理可以通过抑制无氧呼吸,减少乙醇发酵,延缓笋体腐烂。
竹笋采后呼吸作用方式和途径取决于其环境氧含量,符合竹笋生长发育特点。
在空气处理下,竹笋老化过程以无氧呼吸为主、有氧呼吸为辅,有氧呼吸代谢,以EMP、TCA循环和PPP 交替变化进行,0~2天以PPP为主,2~4天以EMP和TCA循环代谢为主,4~6天3种途径几乎停止;
在高氧处理下,竹笋以有氧呼吸为主,无氧呼吸为辅,TCA循环和PPP代谢受到促进,无氧呼吸受到抑制,高氧处理通过提高呼吸速率和PPP和TCA途径比例以及降低笋体乙醇含量延缓竹笋腐烂速率。




